|
CHAPTER VIII.
THE NERVOUS SYSTEM 0F THE FROG.
The Common Frog | ||
8.
CHAPTER VIII.
THE NERVOUS SYSTEM 0F THE FROG.
 [Description:
Image of page 112.
]
[Description:
Image of page 112.
]
The nervous system
consists of the brain, spinal marrow, and nerves.
The whole consists of a soft, white substance,
ultimately composed of minute threads, termed
nerve-fibres, and minute round bodies
called "ganglionic corpuscles."
The brain is contained in the cavity of the skull, and consists of a rounded mass made up of corpuscles and fibres, and itself contains a cavity which is a remnant of the original canal formed by the upgrowth and overclosure of the walls of the primitive groove of the embryo.
The spinal marrow (as has been said earlier) traverses the canal formed by the successive neural arches of the vertebræ, being directly continuous with the brain, which it, as it were, continues on down the back. Like the brain, it is largely composed of corpuscles, as well as fibres, and itself contains a longitudinal cavity (continuous with that in the brain), which is also the ultimate condition
 [Description:
Image of page 113.
]
[Description:
Image of page 113.
]
The nerves generally (which are made up of fibres) proceed forth from the brain and spinal marrow, which latter are therefore called the central, or (from their position along the dorsal axis of the body), the axial portion of the nervous system.
All the nerves which so proceed together constitute what is called the peripheral, or (because going to the limbs which are appendages of the trunk), the appendicular portion of the nervous system.
From the brain proceed the nerves of special sense: a pair, one on each side, going to the nostrils (1, the olfactory nerves), another pair going to the eyes (2, the optic nerves), and a third pair going to the ears within the skull (3, the auditory nerves). Other nerves go to the tongue and palate, ministering to taste, and again others to the little muscles (orbital muscles), which move the eyeball in various directions, and to different parts of the face.
The nerves which come forth from the spinal marrow are called spinal nerves. They proceed out in pairs (one on each side), and are distributed to the limbs and trunk.
Each nerve consists of fibres, of two sorts proceeding respectively from the ventral (in man anterior), and the dorsal (in man posterior) aspects of the spinal marrow. But these two kinds of fibres are distributed side by side in the ramifications and distributions of each nerve.
The fibres which come ultimately from the dorsal aspect of the spinal marrow are those which carry
 [Description:
Image of page 114.
]
[Description:
Image of page 114.
]
The fibres which come ultimately from the ventral aspect of the spinal marrow, are those which carry an influence outwards, and produce a contraction in the muscles, and are therefore called efferent or motor.
It is the nervous system of the frog, rather than any other set of its organs, which has especially excited interest and attention. It is especially to the relations inter se, of the parts of this system that inquiry has been directed. The relations, that is, of its central or axial portion (the brain and spinal column) to its peripheral or appendicular portion (the nerves of the body and limbs).
In the ever memorable year 1789, Galvani accidentally discovered in the separated legs of certain frogs, prepared for broth, those motions produced by irritation of the exposed great nerve of the thigh, now so familiar to most. This action was long called galvanism, after this observer, not, however, that he was absolutely the first to notice a fact of which he was but a re-discoverer—Swammerdam as long ago as 1658 having observed such motions.
They are generally considered as demonstrating the purely "reflex action" of the nervous system— the responsive action, that is, upon muscles, of nervous centres acted on by external stimuli without the intervention of sensation.
It is affirmed that not only will a decapitated frog endeavour to remove an irritating instrument by means of its hind legs and feet; but that if a caustic
 [Description:
Image of page 115.
]
[Description:
Image of page 115.
]
These, and such experiments, are of course conclusive, if the common assumption be conceded that the brain is the indispensable nervous instrument of sensation.
It is possible, however, that the faculty of sensation may be subserved by the spinal cord without the brain, and if so, all these much vaunted experiments are valueless as regards the truth of pure reflex action, not but that they are of extreme interest, as showing what may be done in lower animals without the intervention of any brain action whatever.
 [Description:
Image of page 116.
]
[Description:
Image of page 116.
]
Mr. G. H. Lewes has long contended against the attribution of sensation to the brain exclusively, and Dr. Bastian has recently supported and enforced similar views.
The latter remarks, in his "Beginnings of Life,"— "instead of accepting the popular view, that the brain is the organ of mind, I believe it would be nearer the truth to look upon the whole nervous system as the organ of mind."
Dr. Bastian here uses the word "mind," not as denoting a rational intellect, but as a generic term equivalent to nervous psychical activity.
It may be remarked in passing that these views of Messrs. Lewes and Bastian closely approximate, as far as they go, to that most rational belief that the soul of every creature is whole and entire in every atom of its bodily structure so long as the latter preserves its integrity and vital activity.
The brain of the frog consists of the same essential parts as does the brain of all the vertebrate animals, including man. In the form and in the proportions of those parts, however, it differs extremely from the higher animals (and above all from man), and resembles the lower forms—the brain of the frog (and of Batrachians generally) offering a much closer resemblance to that of a lizard than to that of a mammal.
The brain of man consists of the following fundamental parts:
-
1. A pair (one on each side) of small rounded
bodies, each connected, by a long stalk, with the
mass of the brain, and each shaped somewhat like a
life-preserver. These are the "olfactory
lobes," and
117from the swollen head of each proceed the delicate nerves of smell.
 [Description:
Image of page 117.
]
[Description:
Image of page 117.
]
- 2. An enormous pair of folded masses, which form the great bulk of the human brain, and are called the cerebral lobes or hemispheres. These are so large and preponderant in man, as to hide every other part of the human brain when that organ is viewed from above.
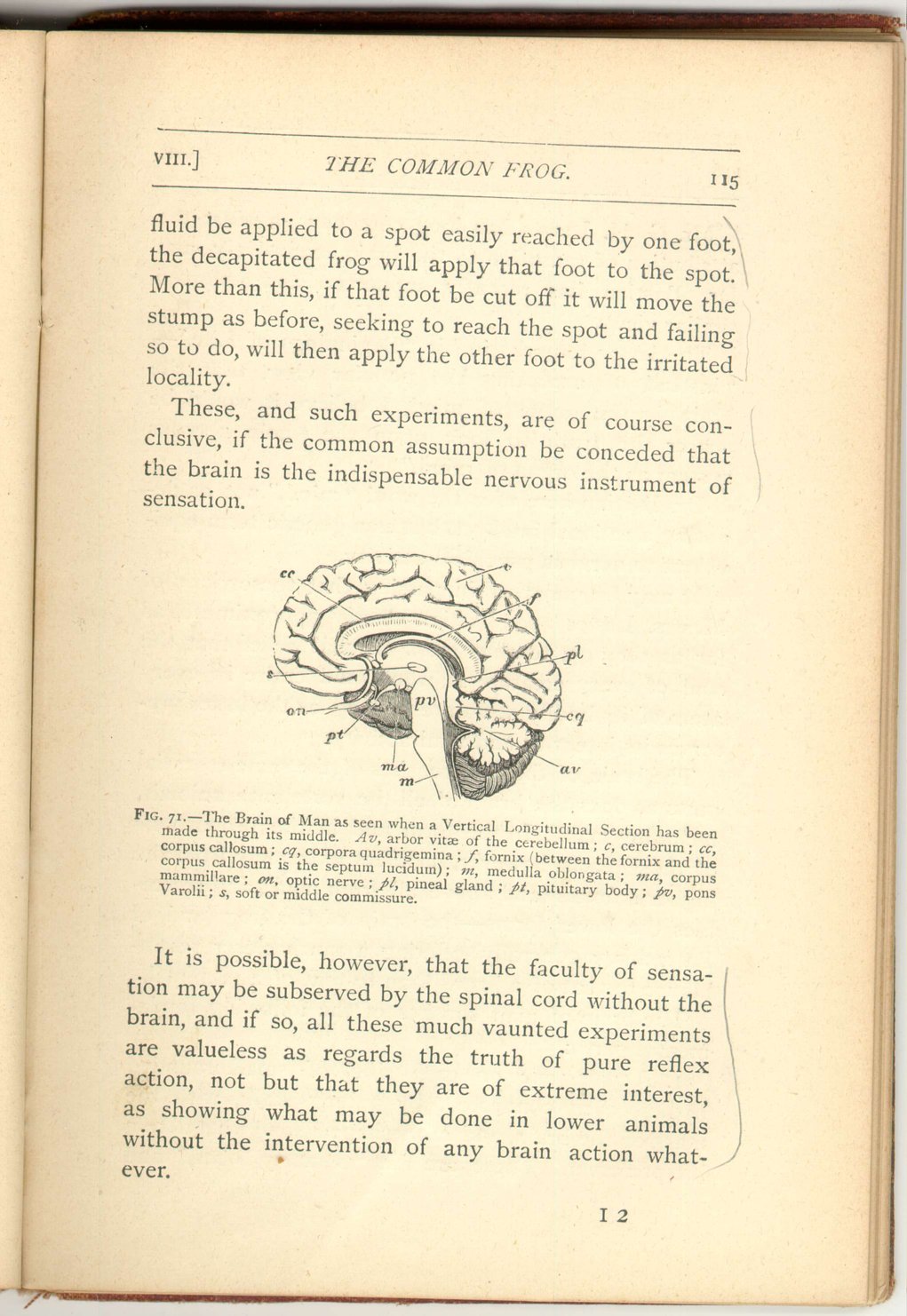
- 3. A relatively very small portion, but one easily recognized, since it supports two conspicuous little bodies. One of these (Figs. 69, 70, 71, pl) is called the pineal gland, and projects more or less upwards; the other (Figs. 69, 70, 71, pt) projects downwards, and is called the pituitary body.
- 4. An also very small portion relatively, is distinguished by bearing certain small prominences (Fig. 69 cq and Fig. 70, na and te) placed behind the pineal gland and called the corpora quadrigemina.
- 5. A rounded mass of finely-folded brain-substance, placed at the lower part of the back of the head, beneath the hinder portion of the cerebral hemispheres. This is called the cerebellum, and when cut through exhibits singular, radiating, tree-like markings, due to the infoldings of the surface of the organ, and called the arbor vitæ (Fig. 70, av).
- 6. That part which directly continues the brain into the spinal marrow (Fig, 71, m). It is overlapped by the cerebellum, and contains that portion of the remnant of the primitive nervous canal, which is named the fourth ventricle. This sixth fundamental part of man's brain is called the medulla oblongata.
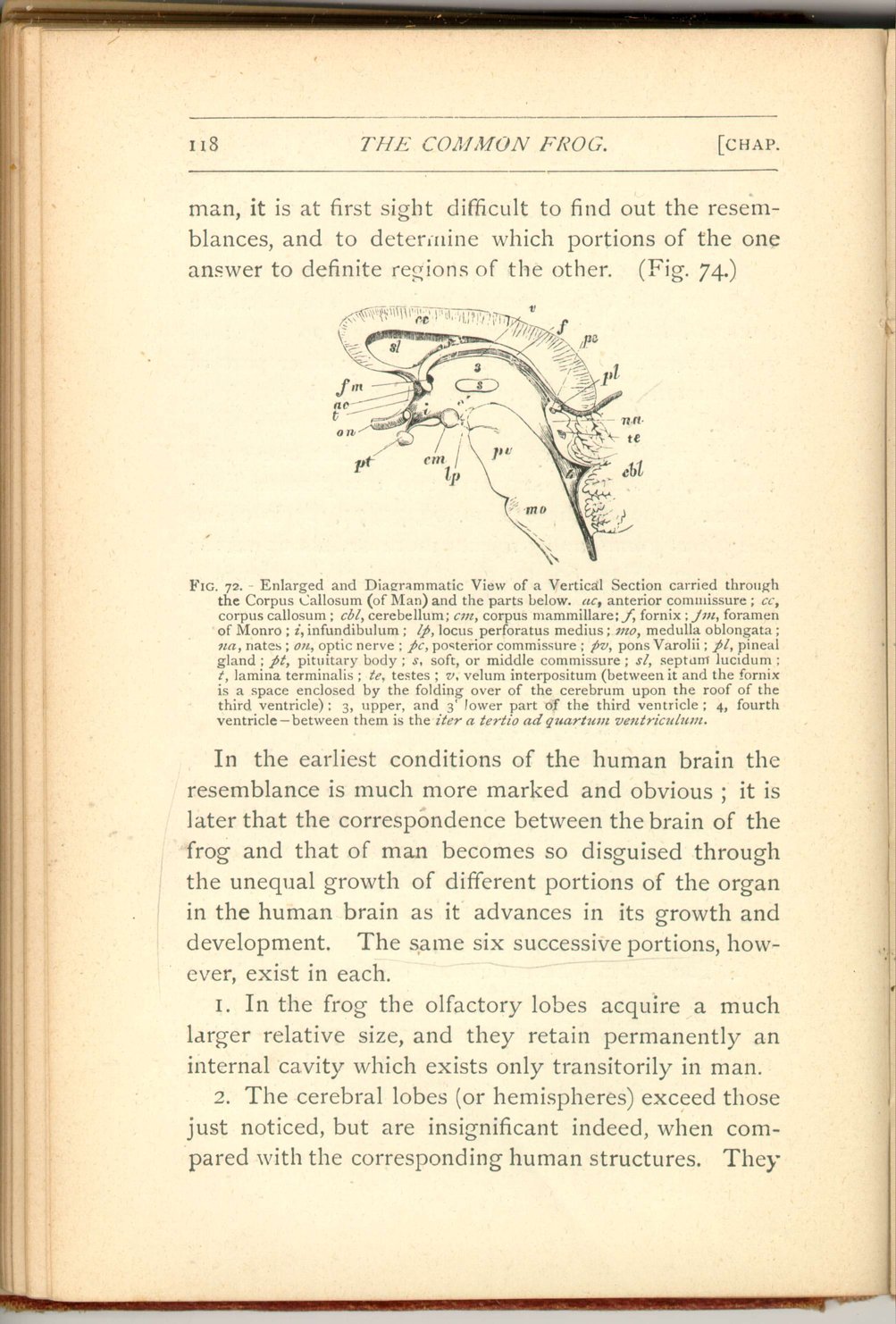
On turning to the brain of the frog from that of
 [Description:
Image of page 118.
]
[Description:
Image of page 118.
]
In the earliest conditions of the human brain the resemblance is much more marked and obvious; it is later that the correspondence between the brain of the frog and that of man becomes so disguised through the unequal growth of different portions of the organ in the human brain as it advances in its growth and development. The same six successive portions, however, exist in each.
- 1. In the frog the olfactory lobes acquire a much larger relative size, and they retain permanently an internal cavity which exists only transitorily in man.
-
2. The cerebral lobes (or hemispheres) exceed those
just noticed, but are insignificant indeed, when
compared with the corresponding human structures. They
119
 [Description:
Image of page 119.
]
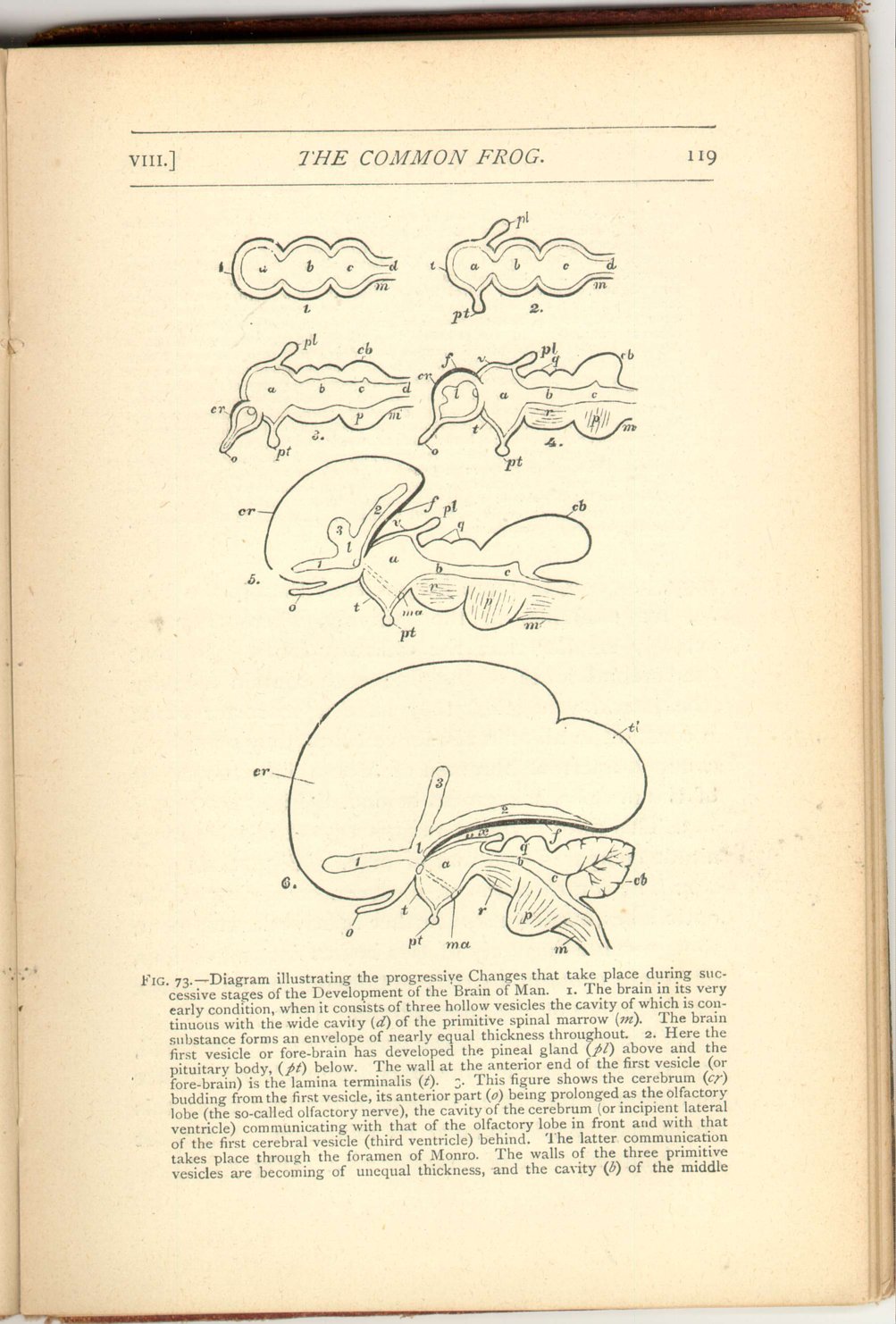
Fig. 73.—Diagram illustrating the progressive Changes that take place during successive stages of the Development of the Brain of Man. 1. The brain in its very early condition, when it consists of three hollow vesicles the cavity of which is continuous with the wide cavity (d) of the primitive spinal marrow (m). The brain substance forms an envelope of nearly equal thickness throughout. 2. Here the first vesicle or fore-brain has developed the pineal gland (pl) above and the pituitary body, (pt) below. The wall at the anterior end of the first vesicle (or fore-brain) is the lamina terminalis (t). 3. This figure shows the cerebrum (cr) budding from the first vesicle, its anterior part (o) being prolonged as the olfactory lobe (the so-called olfactory nerve), the cavity of the cerebrum (or incipient lateral ventricle) communicating with that of the olfactory lobe in front and with that of the first vesicle (third ventricle) behind. The latter communication take place through the foramen of Monro. The walls of the three primitive vesicles are becoming of unequal thickness, and the cavity (b) of the middlemay, however, be more insignificant than in the frog, as, for example, in the lamprey, where they are actually smaller than the olfactory lobes. In that the cerebral lobes of the frog each contain a cavity (the lateral ventricles) they have a character which is constant in all animals above fishes, they open by a common aperture (foramen of Monro) into the cavity of the next brain segment behind. (Fig. 74, b.)120vesicle (iter a tertio ad quartum ventriculum) is becoming reduced in relative size. 4. The cerebrum is here enlarged, and the inequality in thickness of the wall of the primitive vesicle is increased. The thickened upper part of the wall of the cerebrum is the fornix (f). 5. This figure shows the cerebrum still more enlarged, and with a triradiate cavity (l, 1, 2. 3). The fornix has now come to look slightly downwards; dotted lines indicate the downward extension of its anterior part, into the corpora mammillaria. 6. Here the cerebrum is still more enlarged and backwardly extended. The fornix is shown bordering the descending cornu and extending into the temporal lobe (tl) of the cerebrum, which lobe is destined to descend (when the brain is fully developed) so much more that it comes to advance forwards. The fornix borders the margin of the very thin outer wall of the descending cornu, which when torn forms the fissure of Bichat. The bending back of the cerebrum has now almost enclosed (between the fornix and the velum) the space (x) which in Fig. 4 is widely open, making what is morphologically called the outside of the brain come practically to be in its very centre. a, fore-brain; b, mid-brain; c, hind-brain; cb, cerebellum; cr, cerebrum; d, cavity of the medulla; f, fornix; l, lateral ventricle; m, medulla oblongata; ma, corpora mammillaria; o, olfactory lobe; p, pons Varolii; pl, pineal gland; pt, pituitary body; q, corpora quadrigemina; r, crura cerebri; t, lamina terminalis; tl, temporal lobe of the c rebrum; x, space, enclosed by the extension backwards of the cerebrum; 1, anterior cornu of lateral ventricle; 2, its middle or descending cornu; 3, its posterior cornu.
[Description:
Image of page 119.
]
Fig. 73.—Diagram illustrating the progressive Changes that take place during successive stages of the Development of the Brain of Man. 1. The brain in its very early condition, when it consists of three hollow vesicles the cavity of which is continuous with the wide cavity (d) of the primitive spinal marrow (m). The brain substance forms an envelope of nearly equal thickness throughout. 2. Here the first vesicle or fore-brain has developed the pineal gland (pl) above and the pituitary body, (pt) below. The wall at the anterior end of the first vesicle (or fore-brain) is the lamina terminalis (t). 3. This figure shows the cerebrum (cr) budding from the first vesicle, its anterior part (o) being prolonged as the olfactory lobe (the so-called olfactory nerve), the cavity of the cerebrum (or incipient lateral ventricle) communicating with that of the olfactory lobe in front and with that of the first vesicle (third ventricle) behind. The latter communication take place through the foramen of Monro. The walls of the three primitive vesicles are becoming of unequal thickness, and the cavity (b) of the middlemay, however, be more insignificant than in the frog, as, for example, in the lamprey, where they are actually smaller than the olfactory lobes. In that the cerebral lobes of the frog each contain a cavity (the lateral ventricles) they have a character which is constant in all animals above fishes, they open by a common aperture (foramen of Monro) into the cavity of the next brain segment behind. (Fig. 74, b.)120vesicle (iter a tertio ad quartum ventriculum) is becoming reduced in relative size. 4. The cerebrum is here enlarged, and the inequality in thickness of the wall of the primitive vesicle is increased. The thickened upper part of the wall of the cerebrum is the fornix (f). 5. This figure shows the cerebrum still more enlarged, and with a triradiate cavity (l, 1, 2. 3). The fornix has now come to look slightly downwards; dotted lines indicate the downward extension of its anterior part, into the corpora mammillaria. 6. Here the cerebrum is still more enlarged and backwardly extended. The fornix is shown bordering the descending cornu and extending into the temporal lobe (tl) of the cerebrum, which lobe is destined to descend (when the brain is fully developed) so much more that it comes to advance forwards. The fornix borders the margin of the very thin outer wall of the descending cornu, which when torn forms the fissure of Bichat. The bending back of the cerebrum has now almost enclosed (between the fornix and the velum) the space (x) which in Fig. 4 is widely open, making what is morphologically called the outside of the brain come practically to be in its very centre. a, fore-brain; b, mid-brain; c, hind-brain; cb, cerebellum; cr, cerebrum; d, cavity of the medulla; f, fornix; l, lateral ventricle; m, medulla oblongata; ma, corpora mammillaria; o, olfactory lobe; p, pons Varolii; pl, pineal gland; pt, pituitary body; q, corpora quadrigemina; r, crura cerebri; t, lamina terminalis; tl, temporal lobe of the c rebrum; x, space, enclosed by the extension backwards of the cerebrum; 1, anterior cornu of lateral ventricle; 2, its middle or descending cornu; 3, its posterior cornu. [Description:
Image of page 120.
]
[Description:
Image of page 120.
]
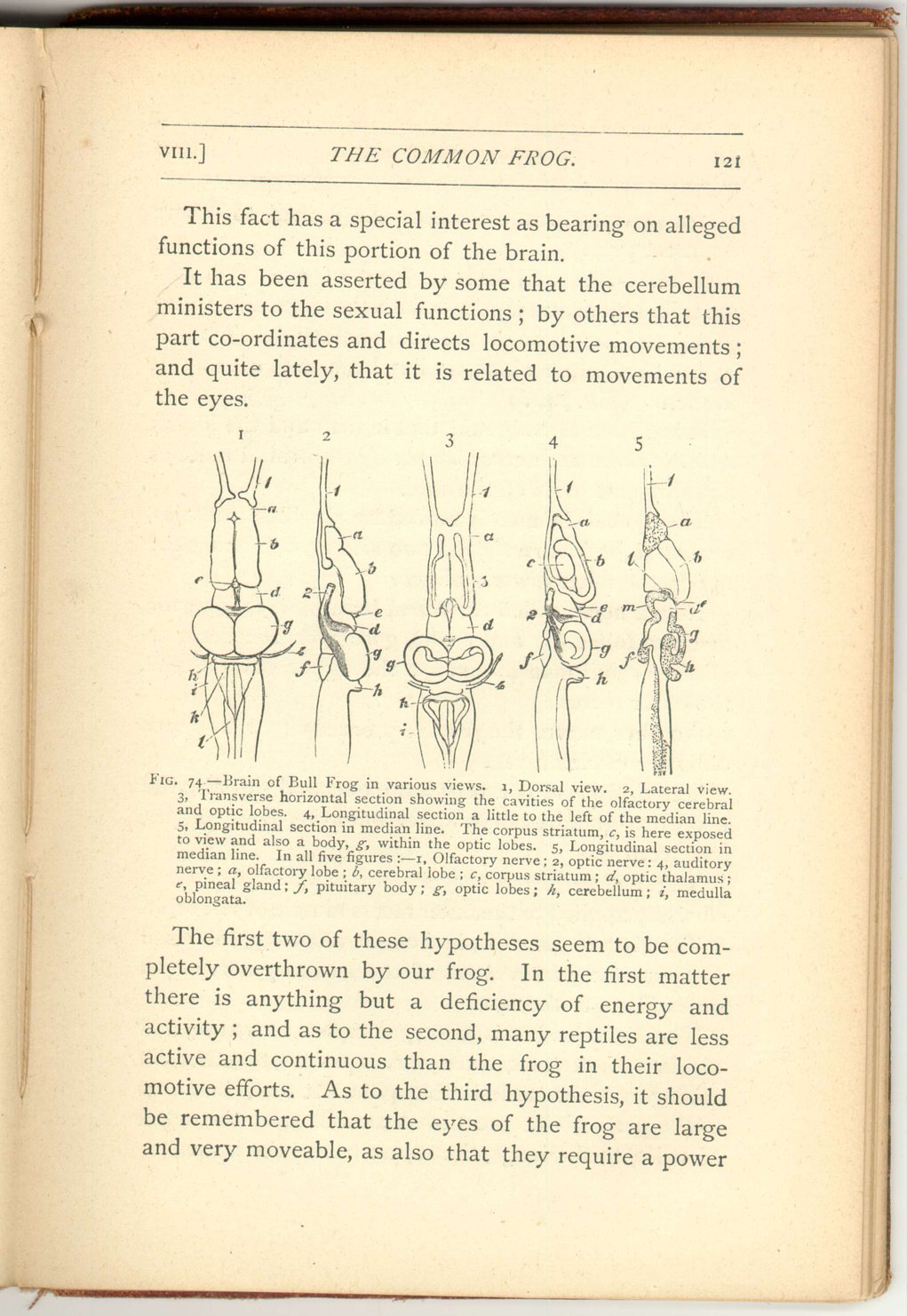
- 3. This third segment retains a great relative magnitude compared with that of man. (Fig. 74, d.)
- 4. The fourth segment, however, consisting of the optic lobes, attains a still further relative development, though consisting only of two bodies instead of four, but these contain a cavity not found in the corpora quadrigemina of the human brain. (Fig. 74, g.)
- 5. The fifth segment, the cerebellum, is very small, and smaller than the same part in animals both higher and lower in the scale; indeed, in the frog class, this organ may be said to be at its minimum. When cut it exhibits no trace of an arbor vitæ. (Fig. 74, h.)
 [Description:
Image of page 121.
]
[Description:
Image of page 121.
]
This fact has a special interest as bearing on alleged functions of this portion of the brain.
It has been asserted by some that the cerebellum ministers to the sexual functions; by others that this part co-ordinates and directs locomotive movements; and quite lately, that it is related to movements of the eyes.
The first two of these hypotheses seem to be completely overthrown by our frog. In the first matter there is anything but a deficiency of energy and activity; and as to the second, many reptiles are less active and continuous than the frog in their locomotive efforts. As to the third hypothesis, it should be remembered that the eyes of the frog are large and very moveable, as also that they require a power
 [Description:
Image of page 122.
]
[Description:
Image of page 122.
]
- 6. The sixth and last segment of the brain, the medulla oblongata, is also relatively large, and is exposed to view through the rudimentary development of the cerebellum which, as has been said, overlaps it in man. (Fig. 74, i.)
It has been already said, that in man and the higher animals there are nerves supplying the orbital muscles and different parts of the face.
The eyeball in man is moved by six little muscles, four straight (the recti) and two oblique, one being the upper, the other lower, oblique.
Now a nerve called the third, because it follows the first two (olfactory and optic), goes from the brain to all the orbital muscles except the upper oblique and the outer rectus.
Another nerve, the fourth, proceeds to the upper oblique muscle only.
The fifth nerve is a very large one, and supplies the nose, tear-gland, eyelids, upper and lower jaws, tongue, and teeth.
The sixth nerve is a very small one indeed, being exclusively applied to the outer rectus muscle of the orbit.
The seventh nerve is, in human anatomy, reckoned as part (the portio dura) of the auditory nerve. It sends fibres to the face.
The eighth nerve is a very complex structure, and consists of, at least, three nerves united together, all arising from the medulla oblongata. It sends branches to the parts about the throat as well as to the organ of voice, to the lungs, the stomach, and the heart.
 [Description:
Image of page 123.
]
[Description:
Image of page 123.
]
The nerves of the frog exhibit certain intermediate conditions, like those we have seen to exist in various other parts of its anatomy.
In the higher vertebrate animals, as in Man, the muscles which move the eyeball are supplied by three distinct nerves, termed respectively the 3rd, 4th and 6th. The 5th nerve being a very large and complex one, sending branches to various parts of the head and its organs.
Now in the frog there is no distinct 6th nerve, it being replaced by an extra branch of the 5th nerve. This modification, however, is but one step towards a condition which obtains in the Mud-fish ( Lepidosiren) when all these three nerves are quite blended with one division (the Ophthalmic) of the 5th nerve.
Again in the higher Vertebrates, as in Man, the 8th nerve is a very large and complex one, and is
 [Description:
Image of page 124.
]
[Description:
Image of page 124.
]
In the tadpole, however, this nerve shows a very different arrangement. After issuing from the skull this nerve sends a branch down the outer side of each branchial arch and then gives off a very long one, which extends laterally, i.e. along the side of the body and tail.
Nothing like this exists in any Beast, Bird, or Reptile, but when we come to the class of Fishes we encounter a precisely similar state of things. Here we find the 8th nerve sending a branch to each branchial arch, and giving off a great nerve proceeding along the side of the body and tail, and on that account named the nervus lateralis.
The eye of the frog is a beautiful and brilliant object, and relatively large. It is furnished with two eyelids, but, unlike those of man, it is the inferior one which is the more moveable. In addition to these it is defended by a third eyelid, called the nictitating eyelid, which is similar to that one which may be seen (if watched for) so frequently and rapidly to cross the eye of birds, e.g. of a hawk.
This structure, however, is no mark of affinity to birds, as it is one which reappears, when wanted, in widely different forms. Thus we find it in the whale, i.e. in the highest class of the Vertebrate sub-kingdom, and in certain sharks, i.e. in the lowest class of the same.
Eyelids do not exist in all members of the frog's class. Even in its order they are extremely minute, in Pipa and Dactylethra, which have very small eyes.
 [Description:
Image of page 125.
]
[Description:
Image of page 125.
]
The ear of the frog's class presents us with the incipient condition of that part as an organ destined to respond to sonorous vibrations conveyed to it by the atmosphere.
In man the internal ear (enclosed in the densest bone of the skull, named, from its density, "petrous") is a very complex organ. The aperture, surrounded by the folds of the external ear, leads by a canal towards a cavity called the tympanic cavity, which cavity is shut off from the exterior by the tympanic membrane (or drum of the ear), which stretches across the canal at a considerable distance from its external aperture. On the inner side of the tympanic cavity lie the convoluted tubes (richly supplied with nerves) which constitute the real organ of hearing or internal ear.
Although the tympanic cavity is shut off from the exterior by the tympanum, it nevertheless is not altogether shut off from the exterior, since it communicates with the back of the mouth by a long and narrow canal termed the Eustachian tube.
It is the existence of these Eustachian openings into the ear from the mouth which causes people when intently listening to keep their mouth slightly open.
In the frog there is no such external canal, but the tympanum is plainly to be seen in the way already described, on the side of the head, covered only by a
 [Description:
Image of page 126.
]
[Description:
Image of page 126.
]
This condition of things, however, does not exist in all the members of the frog's order, still less of his class. But in Proteus, Siren, and Menobranchus there is no tympanic cavity whatever, and the organ of hearing is simply imbedded in the skull, and probably responds but to sonorous vibrations conveyed to it by the denser aquatic medium, and not at all, or but very imperfectly, to those of the atmosphere.
In the ordinary efts we still meet with an Eustachian canal, but the tympanum is absent.
In the frog's own order, as in Pelotates and Bombinator, we may fail to find any tympanum, while the Eustachian tubes are all but obliterated, being reduced to the most minute dimensions.
Another condition, however, may be presented which offers a singular contrast, and is the more remarkable from the widely separated geographical of the forms which present it. In the South American Pipa, as well as in the South African Dactylethra, the two Eustachian tubes run together and open at the back of the mouth, by a median and common aperture.
Strange to say, this is the very condition which exists in birds, though most certainly it cannot be taken as any sign of affinity. In the crocodile these tubes have also a common median opening, but, unlike birds, each tube has also its own lateral opening
 [Description:
Image of page 127.
]
[Description:
Image of page 127.
]
Can the resemblance between Pipa and Dactylethra in this matter be taken as a serious indication of genetic affinity, in spite of the wide, deep, and probably ancient Atlantic which rolls between the two species now?
This is a question which cannot be confidently answered, seeing in how many, other instances structural peculiarities have evidently had an independent origin. Nevertheless, the fact that these two genera agree also in the small size of the eyes, rudimentary eyelids, and vastly expanded sacral transverse process would seem to point to some ancestral and fundamental relationship. If so, however, it is remarkable that no other such forms, or no intermediate ones should have been preserved, seeing that neither kind can be suspected of having migrated to its own habitat from the existing habitat of the other; and therefore that forms similar to that from which we may, if we please, conceive both to have been derived must have had a more or less widely extended geographical distribution and have been numerous in order to have given origin to genera in many respects so different as the two in question.
|
CHAPTER VIII.
THE NERVOUS SYSTEM 0F THE FROG.
The Common Frog | ||