|
CHAPTER VI.
THE SKELETON OF THE FROG
The Common Frog | ||
6.
CHAPTER VI.
THE SKELETON OF THE FROG
 [Description:
Image of page 60.
]
[Description:
Image of page 60.
]
It may cause surprise to
speak of the skin of the common Frog as part of its
skeleton, consisting as the skin does of small membranous
structures only.
The term "skeleton," however, should properly include all the membranous and gristly, as well as the bony structures. [14]
Moreover, more or less of the skin may attain to so solid a condition as to justify its comprehension under the name "skeleton," even in the popular signification of that term.
The skin of Vertebrate animals consists of two Layers: an outer layer (the epidermis or ecteron), and an inner layer (the dermis or enderon). The epidermis, and any projections or processes developed from it when they take on a dense or hardened structure, become horny. Of such horny nature are hairs, feathers, nails, and scales; they are more or less epidermal appendages. The dermis when hardened becomes bony, and of such nature are the bony skin-plates or "scutes" and teeth. They are dermal
 [Description:
Image of page 61.
]
[Description:
Image of page 61.
]
 [Description:
Image of page 62.
]
[Description:
Image of page 62.
]
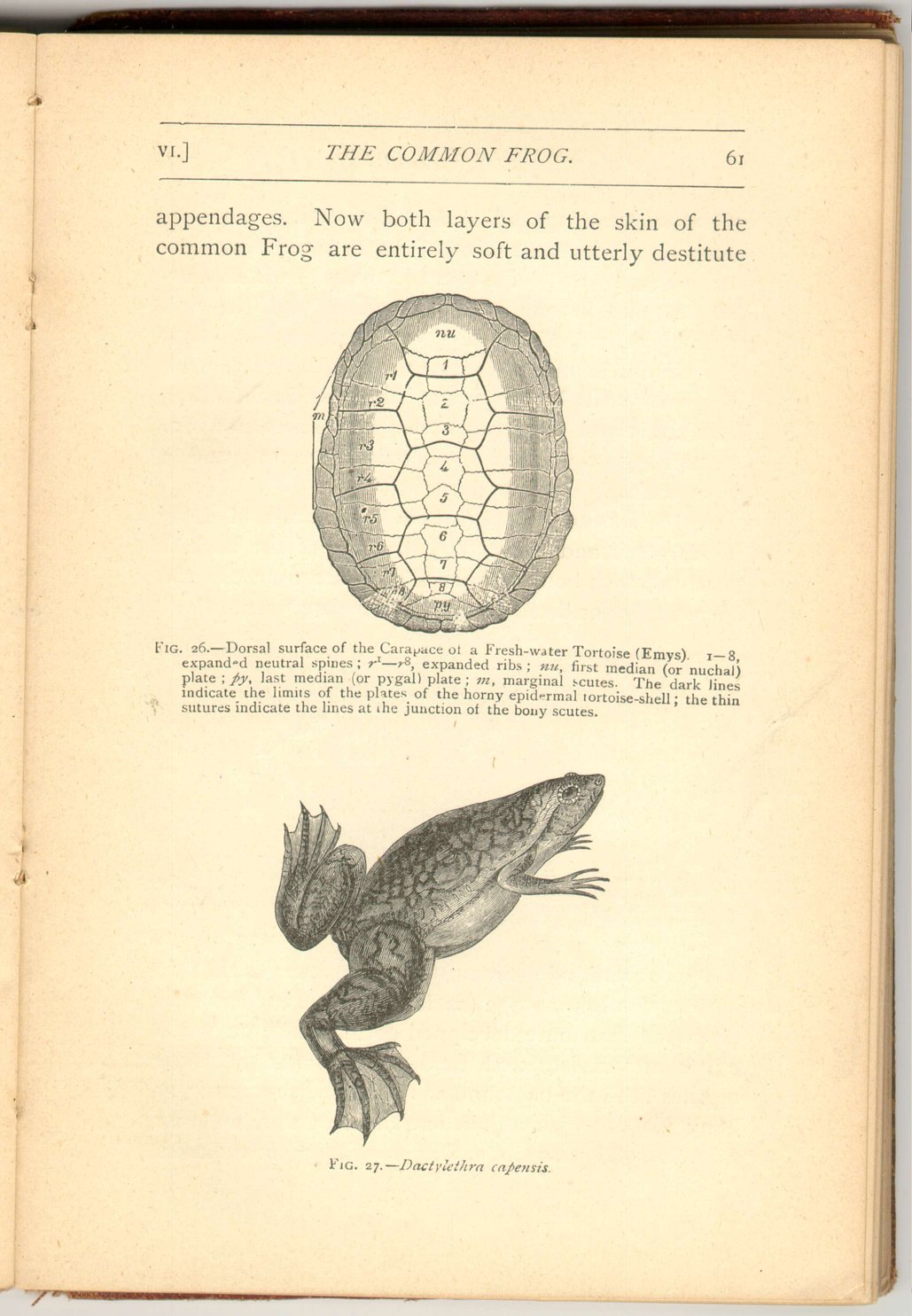
As has been mentioned, in two kinds of Frog (Ceratophrys and Ephippifer) the skin of the back is furnished with bony plates. These are found in the deeper layer or dermis, and are therefore "scutes."
The remarkable circumstance, however, is that we have here a lower stage (as if it were an incipient condition) of that more developed dermal skeleton which exists in tortoises and turtles. In most of these reptiles both the back and the belly are protected by bony plates which adjoin one another, and together form a solid box in which the body is enclosed.
 [Description:
Image of page 63.
]
[Description:
Image of page 63.
]
Again, the middle series of bony plates of the back are continuous with the subjacent joints of the back-bones, and the lateral series of dorsal plates are continuous with the ribs beneath them (fig. 28 ).
There are certain Chelonians, however—"mud-tortoises"—(of the genus Trionyx), which have the dorsal plates much less developed and not connected
 [Description:
Image of page 64.
]
[Description:
Image of page 64.
]
Have we then here a true sign of genetic affinity? Are these tortoises to be deemed the more specially modified, descendants of shielded frogs or of some, as yet unknown, slightly-shielded animals which were the common ancestors both of frogs and tortoises?
Certainly tortoises cannot be the direct descendants of frogs, they agree with all reptiles in characters which are both too numerous and too important to allow such an opinion to be entertained for a moment.
The other opinion is hardly less untenable; for if all the multitudinous species of frogs (together with a number of reptilian forms more closely allied to the tortoise than any frogs are) descended from slightly shielded animals, how comes it that all frogs and toads, save one or two species in no other way peculiar, have every one of them lost every trace of such shielded structure, which nevertheless cannot easily be conceived to have been in any way prejudicial to their existence and survival?
On the other band, it cannot but strike us with surprise that structures so similar—extending even to
 [Description:
Image of page 65.
]
[Description:
Image of page 65.
]
The skin of the frog is also interesting from a physiological point of view. Our own skin is by no means popularly credited with the great importance really due to it. "Only the skin!" is an exclamation not unfrequently heard, and wonder is very often felt when death supervenes after a burn which has injured but a comparatively small surface of the body. Yet our skin is really one of our most important organs, and is able to supplement, and to a very slight extent even to replace, the respective actions of the kidneys, the liver, and the lungs. [15]
In the frog we have this cutaneous activity developed in a much higher degree. Not only does its perspiratory action take place to such an extreme degree that a frog tied where it cannot escape the rays of a summer's sun speedily dies—nay, more, is soon perfectly dried up—but, its respiratory action is both constant and important. This has been experimentally demonstrated by the detection of the carbonic acid given out in water by a frog over the head of which a bladder had been so tightly tied as to prevent the possibility of the escape of any exhalation from the lungs. The fact of cutaneous respiration
 [Description:
Image of page 66.
]
[Description:
Image of page 66.
]
It is no less true that in Batrachians which breathe by means of permanent gills—as, e.g. the Axolotl—
 [Description:
Image of page 67.
]
[Description:
Image of page 67.
]
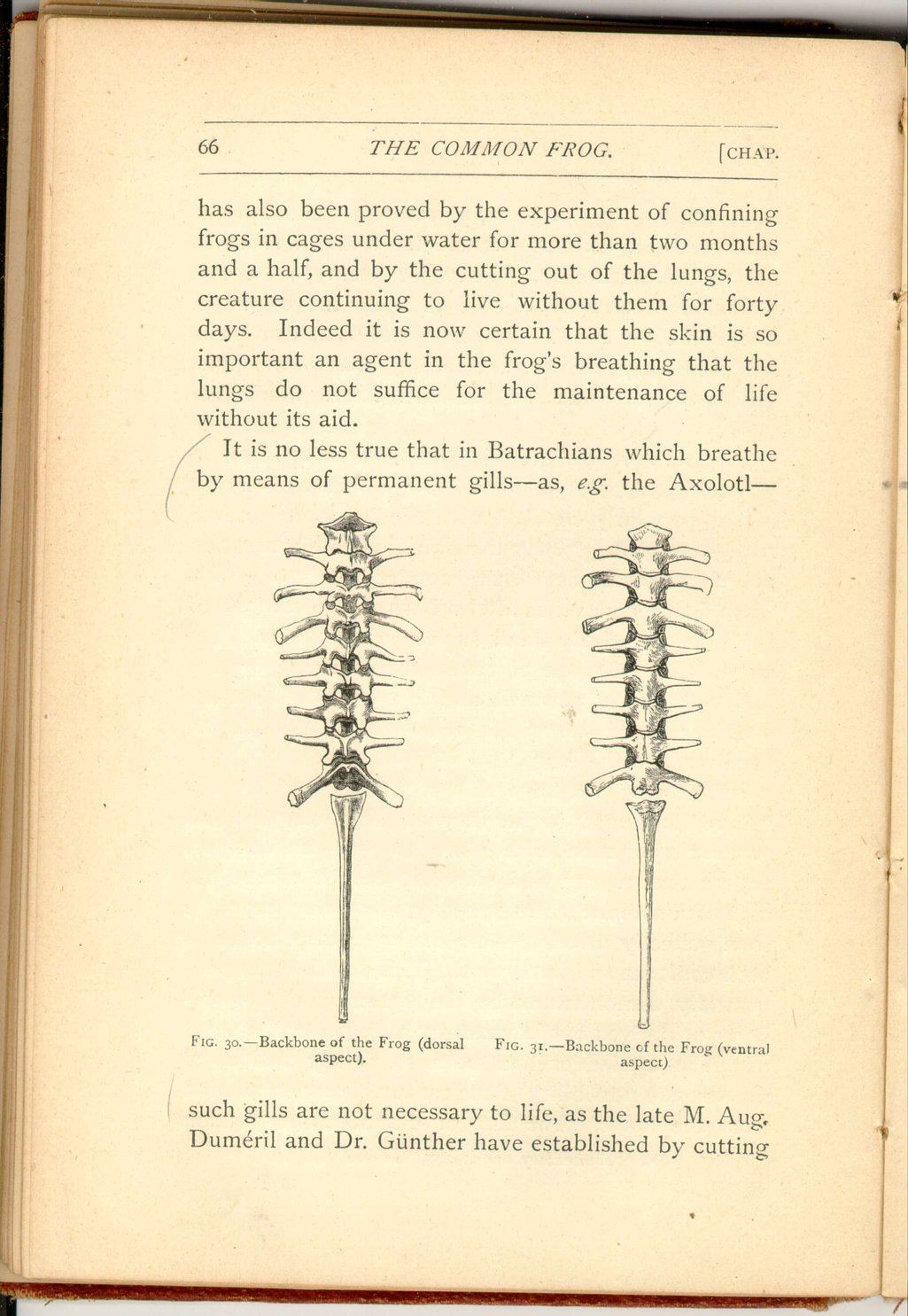
The internal skeleton (or the skeleton commonly so called) of the frog presents some points of considerable interest, especially as exhibiting its intermediate position between fishes on the one hand, and higher vertebrates on the other. First, as regards the backbone, it may be remembered that it is made up of distinct bony joints (or vertebræ), in which it agrees with all animals above fishes and with bony fishes; its hinder termination, however, is essentially fish-like.
It is fish-like because the terminal piece, as it is called, or "coccyx" (unlike the coccyx in man or in birds) is not composed of rudimentary vertebræ which subsequently blend and anchylose together, but is formed by the ossification continuously of the membrane investing (or sheath of) the hindermost part of that primitive continuous rod, or notochord, [16] which, as has been said, precedes, in all vertebrate animals, the development of the backbone, making its appearance beneath the primitive groove.
The vertebræ are shaped like rings, and enclose within their circuit the spinal marrow upon which, as it were, these rings are strung. From the side of each
 [Description:
Image of page 68.
]
[Description:
Image of page 68.
]
That part of each vertebra which is placed next the body cavity is generally the thickest part, and is called the "body," or "centrum." The series of
The Frog order is distinguished amongst vertebrates as that which has the absolutely smallest number of joints in the backbone. In the frog there are but nine in the front of the coccyx. In the Pipa toad
 [Description:
Image of page 69.
]
[Description:
Image of page 69.
]
In all higher vertebrates, i.e. in all beasts, birds, and reptiles, the head is supported on an especially ring-like vertebra which—because it so supports—is called the atlas, and this (in almost all) can turn upon a peculiar vertebra termed (from this circumstance)
In the frog (and in all its class) we find but a single vertebra representing these two, but in some allied forms, e.g. in Amphiuma, this vertebra develops a
 [Description:
Image of page 70.
]
[Description:
Image of page 70.
]
The frog, as has been said, has no ribs, in spite of the long "transverse processes" which project out on each side of the backbone. Ribs are not necessary to it, for it could apply them to none of the purposes to which ribs are ever applied.
In all beasts ribs aid importantly in respiration, serving by their motions alternately to inflate or empty the lungs by enlarging or contracting the cavity of the chest in the way before mentioned. The frog, however, breathes exclusively, as regards the lungs, by swallowing air by a mechanism which will be described shortly.
In serpents the ribs are the organs of locomotion, as also in the Flying Dragon before referred to; but in frogs locomotion is effected exclusively by the limbs. In the very aberrant species, Pipa and Dactylethra, there are on each side of the anterior parts of the body two enormously long transverse processes, each process bearing at its extremity a short flattened, straight osseous or cartilaginous rib. These little ribs can, however, take no part in such functions as those just referred to.
Ribs, moreover, are found in the other existing orders of the frog's class, i.e. both in the Urodela and Ophiomorpha. In none, however, do they join a breast-bone, or sternum, another character in which Batrachians agree with fishes, though they differ from fishes in that they have a sternum at all. In ascending from fishes through the Vertebrate sub-kingdom, a sternum first appears in the class Batrachia.
 [Description:
Image of page 71.
]
[Description:
Image of page 71.
]
The several joints of the backbone are connected together by surfaces which are not the same on both the anterior and posterior sides of the centrum, or body, of the same vertebra. Each of the first seven vertebræ is furnished with a round prominence, or head, on the hinder side of its centrum, and each of the precoccygeal vertebra, except the first and last, has the anterior surface of its centrum excavated as a cup for the reception of the ball of the hinder surface of the vertebra next in front. The first vertebra has in front two concavities, side by side, to articulate with the skull. The eighth vertebra has a concavity at each end of its "body." The ninth vertebra has a body provided with a single convexity in front and a double convexity behind, to articulate with the concavities placed side by side on the front end of the coccyx. These arrangements are not constant in the frog's order, still less in its class. In Bombinater and Pipa the vertebra are concave behind each centrum, instead of in front: and the same is the case in Salamandra. In many tailed Batrachians the vertebræ are biconcave, as e.g. in Spelerpes, Amphiuma, Proteus, and Siren.
The biconcave shape is an approximation towards the condition which is almost universal in bony fishes,
 [Description:
Image of page 72.
]
[Description:
Image of page 72.
]
Vertebræ with a cavity in front of the centrum and a ball behind it are found in the crocodiles now living as well as in the frog, while vertebræ with a ball in front and a concavity behind are found even amongst beasts as in the joints of the neck of Ruminants, e.g. the sheep. Thus though the vertebræ of the frog's class exhibit no very decided signs of affinity, they show more resemblance to those of fishes than to those of any other non-batrachian class.
The transverse processes of the ninth or last vertebra in front of the coccyx, articulate with the haunch bones, but are not very remarkable in shape. In some frogs and toads the transverse processes of this vertebra become enormously expanded, and the expanded or non-expanded condition of this part is
 [Description:
Image of page 73.
]
[Description:
Image of page 73.
]
The skull of the frog presents numerous points of interest, but only four of these can be here referred to, as other matters demand our attention.
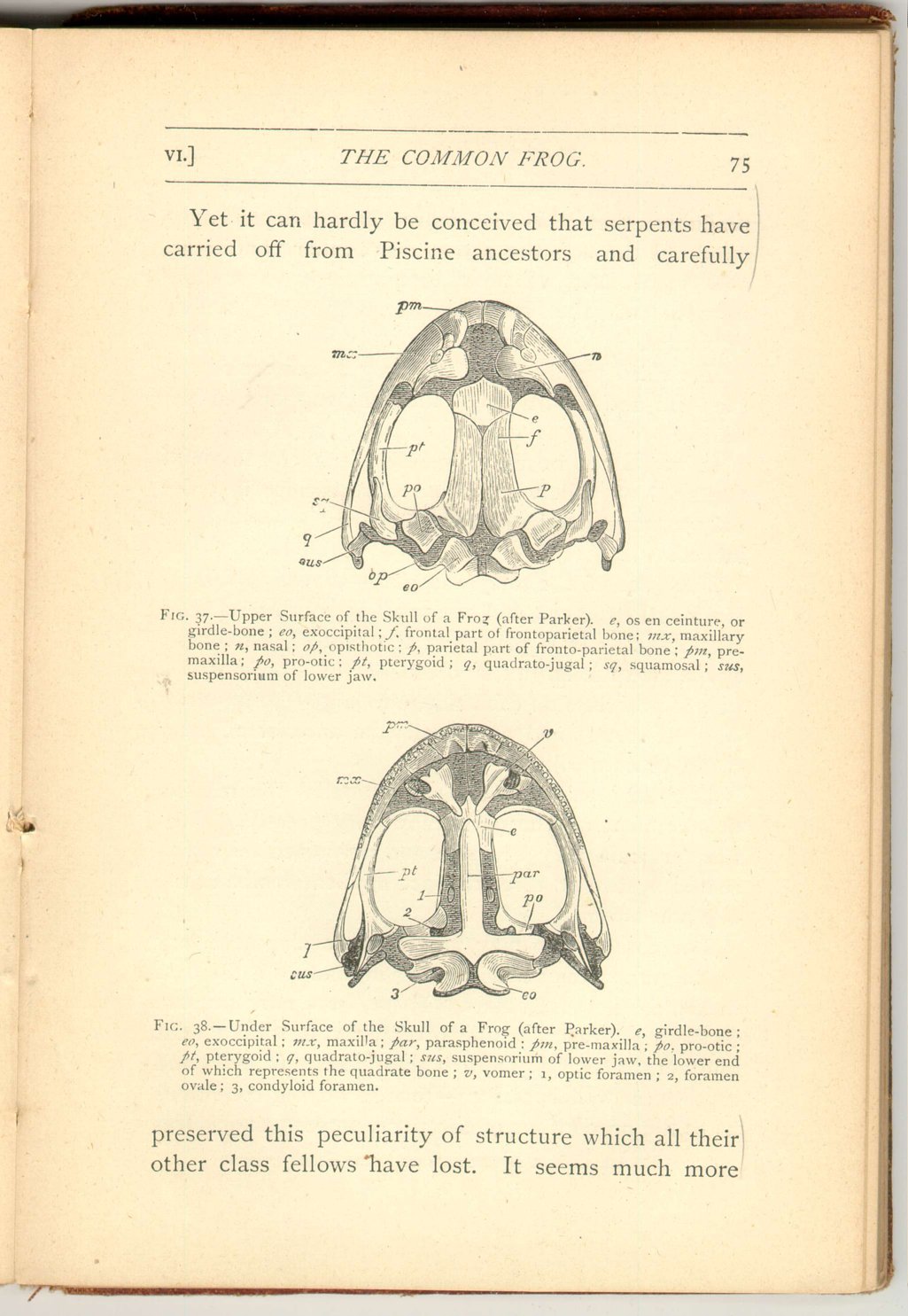
The first of these four relates to its mode of articulation with the vertebral column. As has been said, the first vertebra presents a pair of concavities to the skull. The skull develops from its hinder (or occipital) region a corresponding pair of articular convexities or "condyles." Now in this matter the frog differs from both birds and reptiles, every member of those classes possessing a single median (occipital) condyle for articulation with the vertebral column.
Yet every member of the frog class, not only every toad and newt, but also every species of the Ophiomorpha, and even every one of the long extinct Labyrinthodons (with the doubtful exception of the probably immature and larval Archegosaurus) has a similar pair of occipital condyles. The interesting matter is that man and all beasts have also two occipital condyles. Is this then a mark of affinity,
 [Description:
Image of page 74.
]
[Description:
Image of page 74.
]
We shall presently see that other yet more striking facts may be brought forward in support of the latter view. Nevertheless it must be remembered that there are fishes, though very few and exceptional, which also possess a pair of occipital condyles, and that in one respect most fishes are more like mammals than are any Batrachians, since they, like mammals, have a well-ossified median bone at the base of the skull in the occipital region, a structure which all Batrachians, without a single exception, are destitute of.
The second point of interest concerns the lower part, or base, of the skull, which exhibits a striking agreement with the same part as developed in bony fishes.
This agreement consists in the fact that the middle of the floor of the skull is not formed as in all beasts, birds, and reptiles, by a deposition of bony substance in pre-existing gristle (ossification of cartilage), to which the name Basi-sphenoid is applied, but, as in bony fishes, by a great bone called Parasphenoid, which shoots forwards and also extends backwards to the hinder end of the skull floor, but is formed by the decomposition of bony substance in pre-existing membrane.
Although this great membrane bone is constant in Batrachians and bony fishes, and is represented, if at all, only by minute rudiments in higher vertebrates; nevertheless in serpents we once more meet with a far-reaching and well-developed parasphenoid.
 [Description:
Image of page 75.
]
[Description:
Image of page 75.
]
Yet it can hardly be conceived that serpents have carried off from Piscine ancestors and carefully
 [Description:
Image of page 76.
]
[Description:
Image of page 76.
]
The third peculiarity of the frog's skull consists in the form and conditions of the bony supports of the tongue.
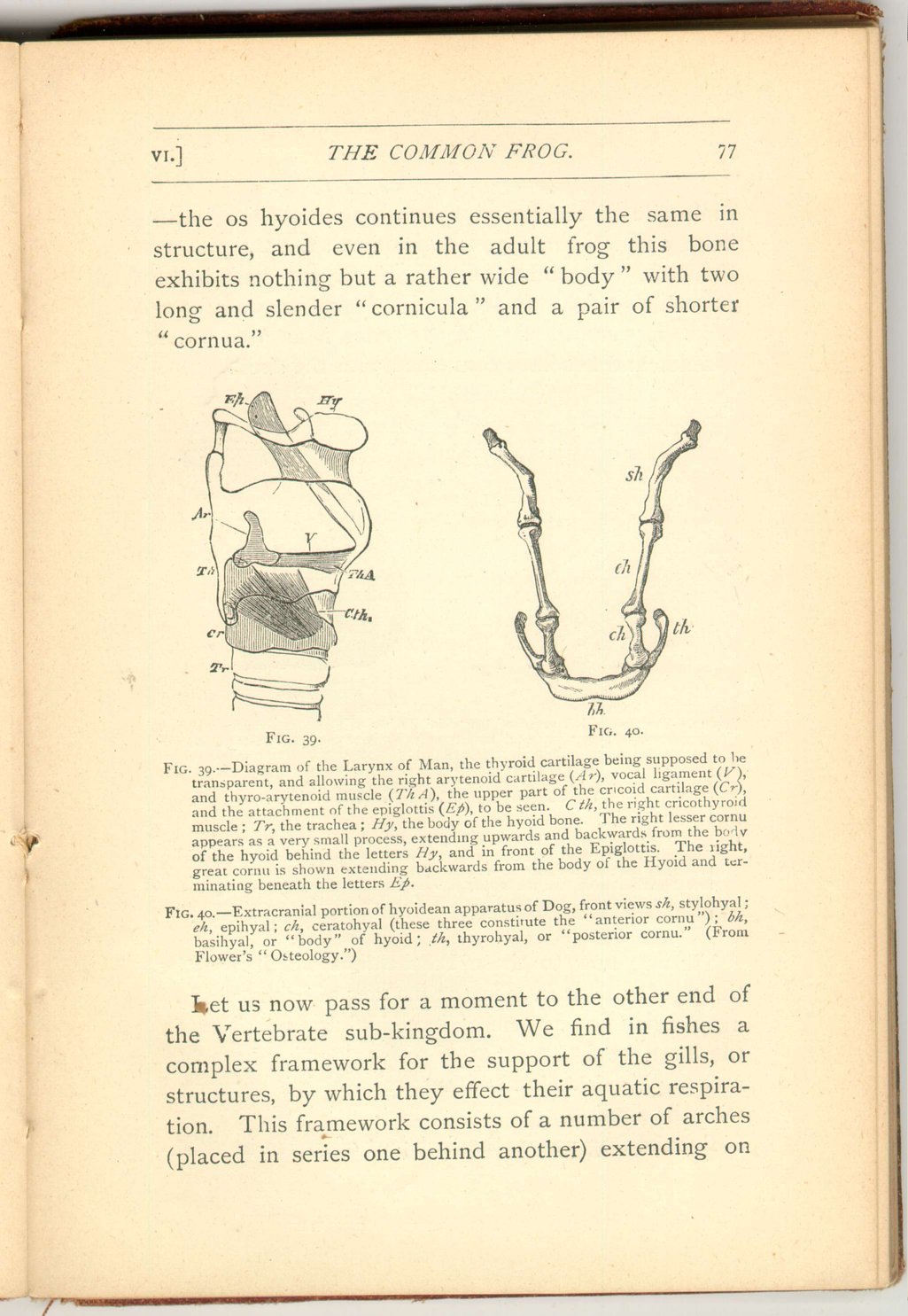
It would not be easy to find a better example of the need of widely extended observations in order duly to understand structures apparently very simple indeed. The bone of the tongue in man—the os hyoides [18]—is a small structure, and one to all appearance of little significance It is placed at the root of the tongue and above the larynx, and consists of a body with a pair of processes on each side, one large (the posterior or great cornu), and one small. (the anterior or lesser cornu, or corniculum).
Even in man's own class (mammalia) the relative development of the parts may vary greatly. Thus the cornicula may be large and may each be represented by two or three distinct applications as in the dog and horse.
The cornua also may take on. a development very much greater than that existing in man as is the case in some other Mammals. These facts may prepare us to expect much greater divergences in lower forms; and yet throughout the two great classes of birds and reptiles (as well as beasts)—though varying conditions as to the proportions of the parts present themselves
 [Description:
Image of page 77.
]
[Description:
Image of page 77.
]
Let us now pass for a moment to the other end of the Vertebrate sub-kingdom. We find in fishes a complex framework for the support of the gills, or structures, by which they effect their aquatic respiration. This framework consists of a number of arches (placed in series one behind another) extending on
 [Description:
Image of page 78.
]
[Description:
Image of page 78.
]
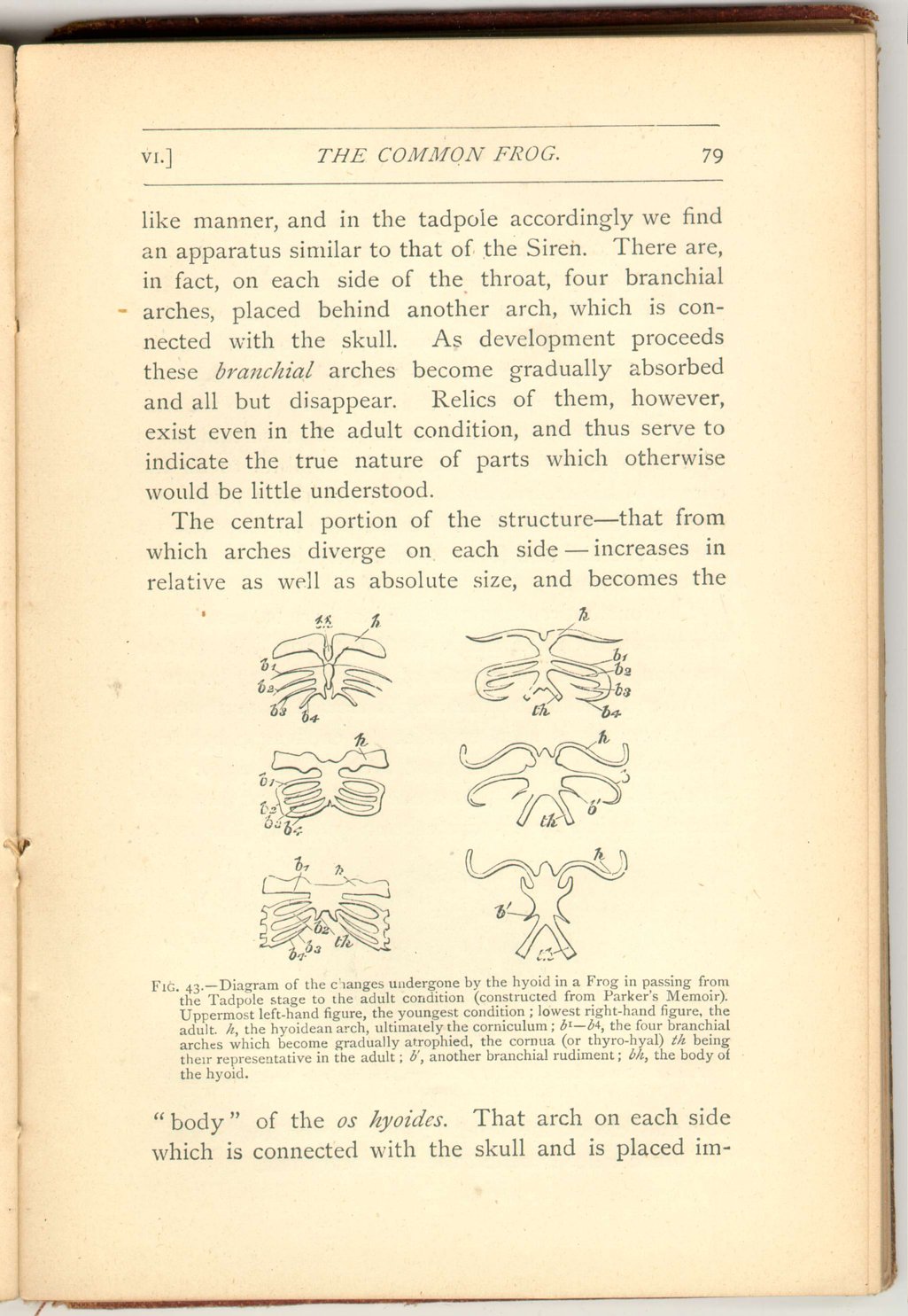
Turning now to those Batrachians which breathe throughout their lives in the manner of fishes, we find a corresponding system of branchial arches. Thus in the Siren we find a series of gill-supporting branchial arches, placed behind another arch which is connected with the skull.
But the frog passes the first part of its life in a fishlike
 [Description:
Image of page 79.
]
[Description:
Image of page 79.
]
 [Description:
Image of page 80.
]
[Description:
Image of page 80.
]
Thus the anatomy of the tongue-bone of the frog, studied in its progressive changes, reveals to us that otherwise unsuspected relations exist in certain parts of the tongue-bone of man. It exhibits to us the cornua of his os hyoides as related to those large and complex branchial arches which play so important a part in the fish and form so relatively large a portion of its skeleton.
The fourth circumstance (the last here to be noticed) connected with the frog's skull concerns the relative position and size of certain of its enveloping bones.
When the skull of the frog is viewed from above, a large vacuity is seen to exist on each side, between the brain-case and the great arch of the upper jaw. In the hinder part of this space is situate the temporal muscle, which by its contraction pulls up the lower
 [Description:
Image of page 81.
]
[Description:
Image of page 81.
]
This very singular structure is found to exist also in the marine turtles, amongst the Chelonians, and here we have another striking resemblance between the Chelonia and the Anoura, apparently reinforcing the argument for the existence of real affinity derived from the presence of such bony dorsal shields in both those two orders. The importance of this character might seem the more unquestionable, since no other reptiles and no birds or beasts whatever were known to exhibit a similar structure.
Quite recently, however, Prof. Alphonse Milne-Edwards has described a beast from Africa (Lophiomys) belonging to the Rodent (rat, rabbit, and squirrel)
 [Description:
Image of page 82.
]
[Description:
Image of page 82.
]
This unexpected discovery completely destroys any weight which might be attached to this character as an evidence of genetic affinity. It does so, because it is inconceivable that this Rodent should have directly descended from a common progenitor of frogs and of Chelonians through a line of ancestors which never lost this cranial shield, though the ancestors of all other beasts, all birds, and all reptiles, except turtles, did lose it. It is inconceivable, for if it were true, a variety of the lowest mammals (Marsupials [19] and Monotremes [20] ) must have less diverged from the ancient common stock than have the members of the Rodent order, and nevertheless these lowest mammals exhibit no trace whatever of such a cranial shield. Here then we have an undoubted example of the
 [Description:
Image of page 83.
]
[Description:
Image of page 83.
]
Here, also, we have a memorable caution against hasty inferences from structural similarities. If this resemblance and that of the dorsal shields are, when taken together, no signs whatever of special genetic affinity—it is difficult to say what structural likenesses are to be deemed unquestionable evidences of a common ancestry.
Passing now to the skeleton of the limbs, we come to a character of great significance, as it is one which serves to distinguish all the limbed species of the frog's class from lower vertebrates. The character is very significant, because all Batrachians, in spite of their numerous and important fish affinities, differ from all fishes, and agree with all higher classes in that they— if they have limbs at all—have them divided into those very typical segments which exist in man; namely, shoulder-bones, arm-bones, wrist-bones, and hand-bones; and into haunch-bones, leg-bones, ankle-bones, and foot-bones respectively. It is difficult, then, to avoid the belief that in the Batrachian class we come
 [Description:
Image of page 84.
]
[Description:
Image of page 84.
]
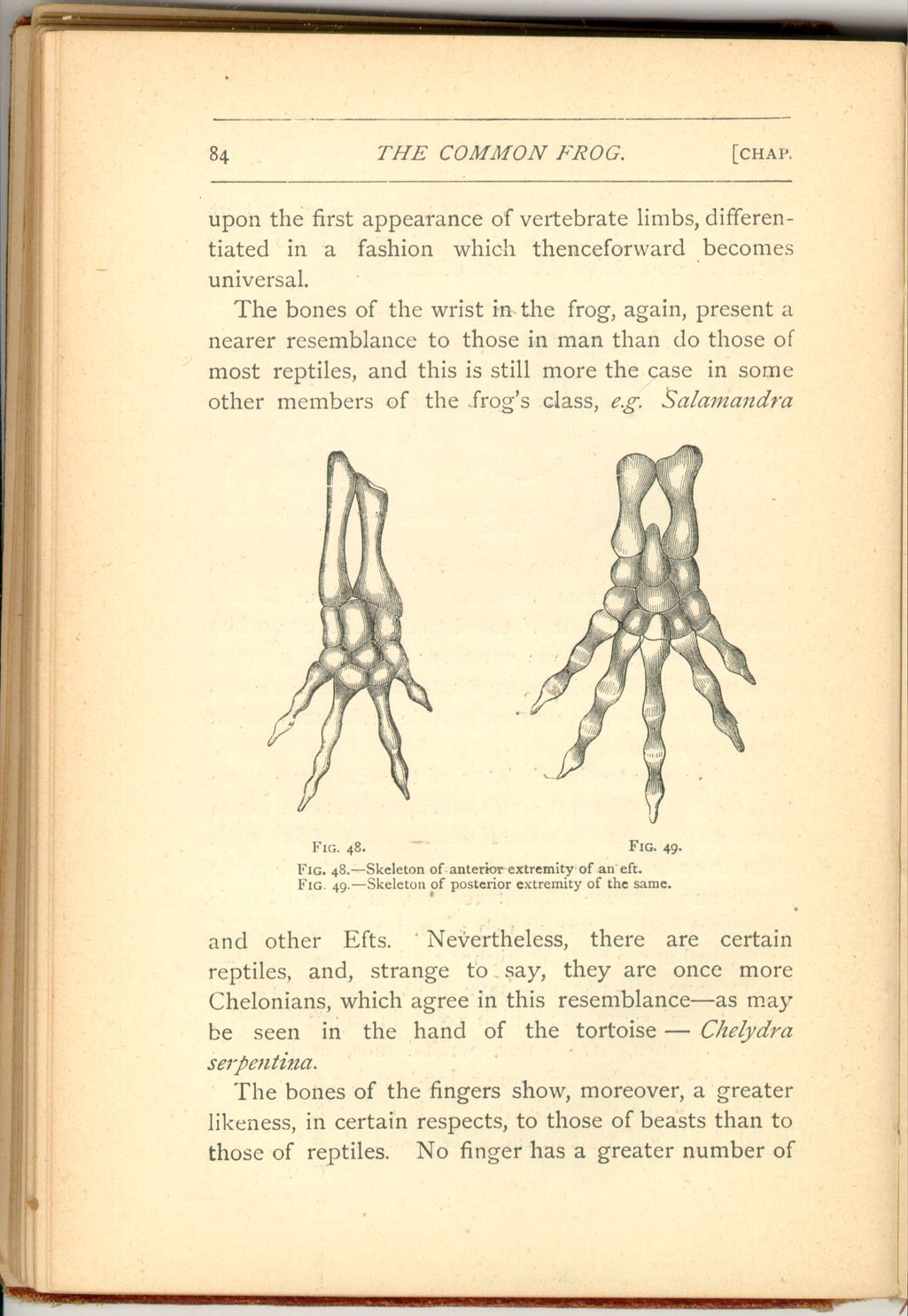
The bones of the wrist in the frog, again, present a nearer resemblance to those in man than do those of most reptiles, and this is still more the case in some other members of the frog's class, e.g. Salamandra
 [Description:
Image of page 85.
]
[Description:
Image of page 85.
]
In the frog the wrist-bones (called respectively the magnum and unciforme) which support the third, fourth, and the little fingers, are formed together into a single ossicle. The same condition, however, sometimes occurs even in the orang. On the other hand, the single bone which in man and beasts supports both the "ring" and the "little" fingers, may be represented by two ossicles in the frog's class (as e.g. in Salamandra, and in some reptiles (as e.g. in Chelydra).
No member of the frog's class which has an arm at all, bears less than two fingers (as in Proteus) upon it. Thus we meet with a number as small as that which is developed amongst beasts in ruminants, but not so small a number as in the horse.
In the rudimentary condition of its thumb the frog participates in a very common defect, since this member is absent in very many forms. It is so even in creatures as highly organised and as like man in bodily structure as monkeys, since both the spider-monkeys
 [Description:
Image of page 86.
]
[Description:
Image of page 86.
]
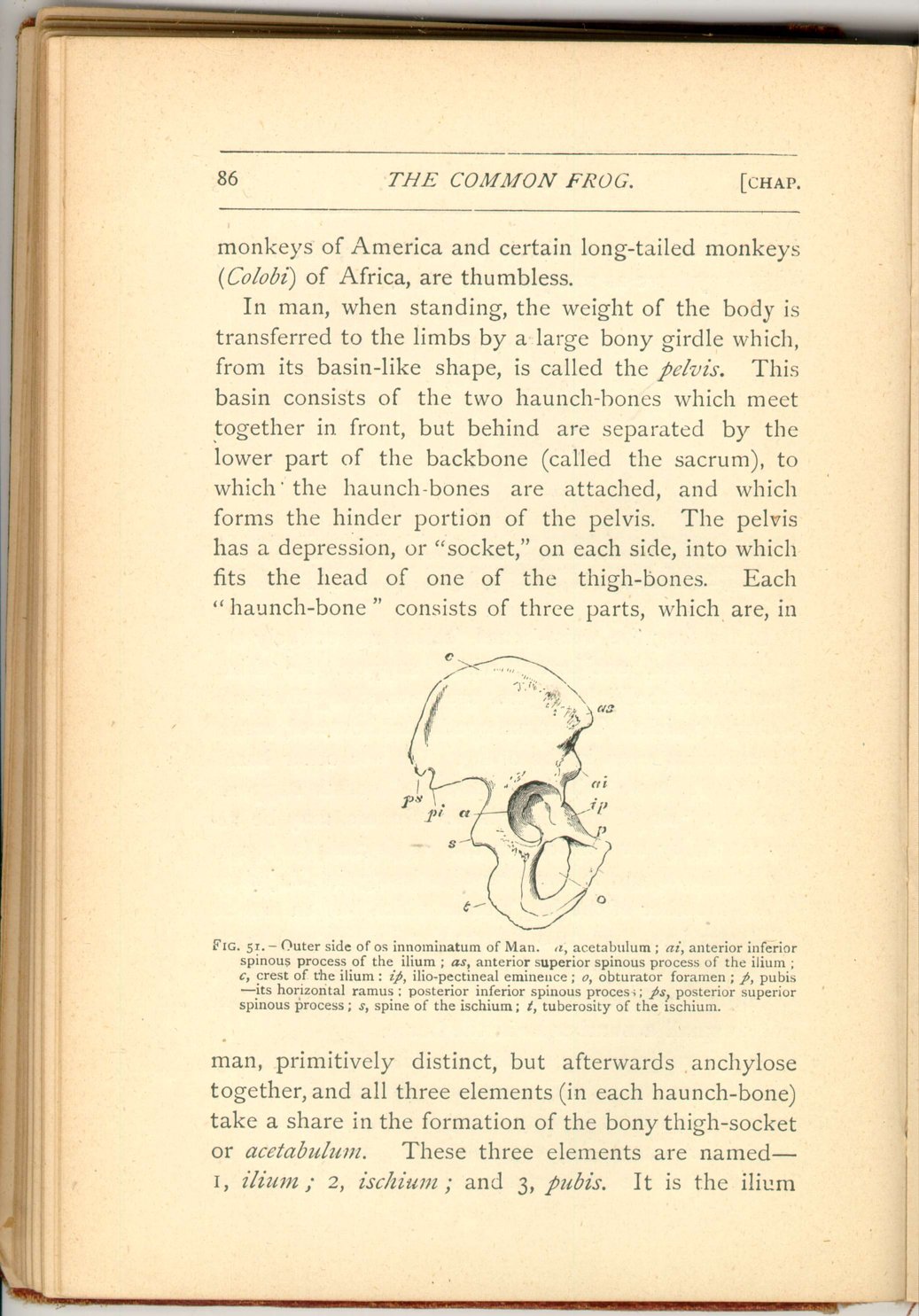
In man, when standing, the weight of the body is transferred to the limbs by a large bony girdle which, from its basin-like shape, is called the pelvis. This basin consists of the two haunch-bones which meet together in front, but behind are separated by the lower part of the backbone (called the sacrum), to which the haunch-bones are attached, and which forms the hinder portion of the pelvis. The pelvis has a depression, or "socket," on each side, into which fits the head of one of the thigh-bones. Each "haunch-bone" consists of three parts, which are, in
 [Description:
Image of page 87.
]
[Description:
Image of page 87.
]
The pelvis of man is often quoted as one of the most peculiar and characteristic parts of his skeleton, and its shape in him is very peculiar. Nevertheless the pelvis as it exists in frogs and toads is a far more exceptional structure. It is so in the extraordinary elongation, yet small vertebral attachment, of the haunch-bones (ilia), as also in the fact that these bones as well as the other pelvic elements (ischia and pubes) are all closely applied to each other in the middle line
 [Description:
Image of page 88.
]
[Description:
Image of page 88.
]
The skeleton of the ankle as developed in the frog's class presents us with some characters, which, more than even those of the wrist, suggest the passage of the line of affinity directly from Batrachians to mammals leaving both reptiles and birds on one side.
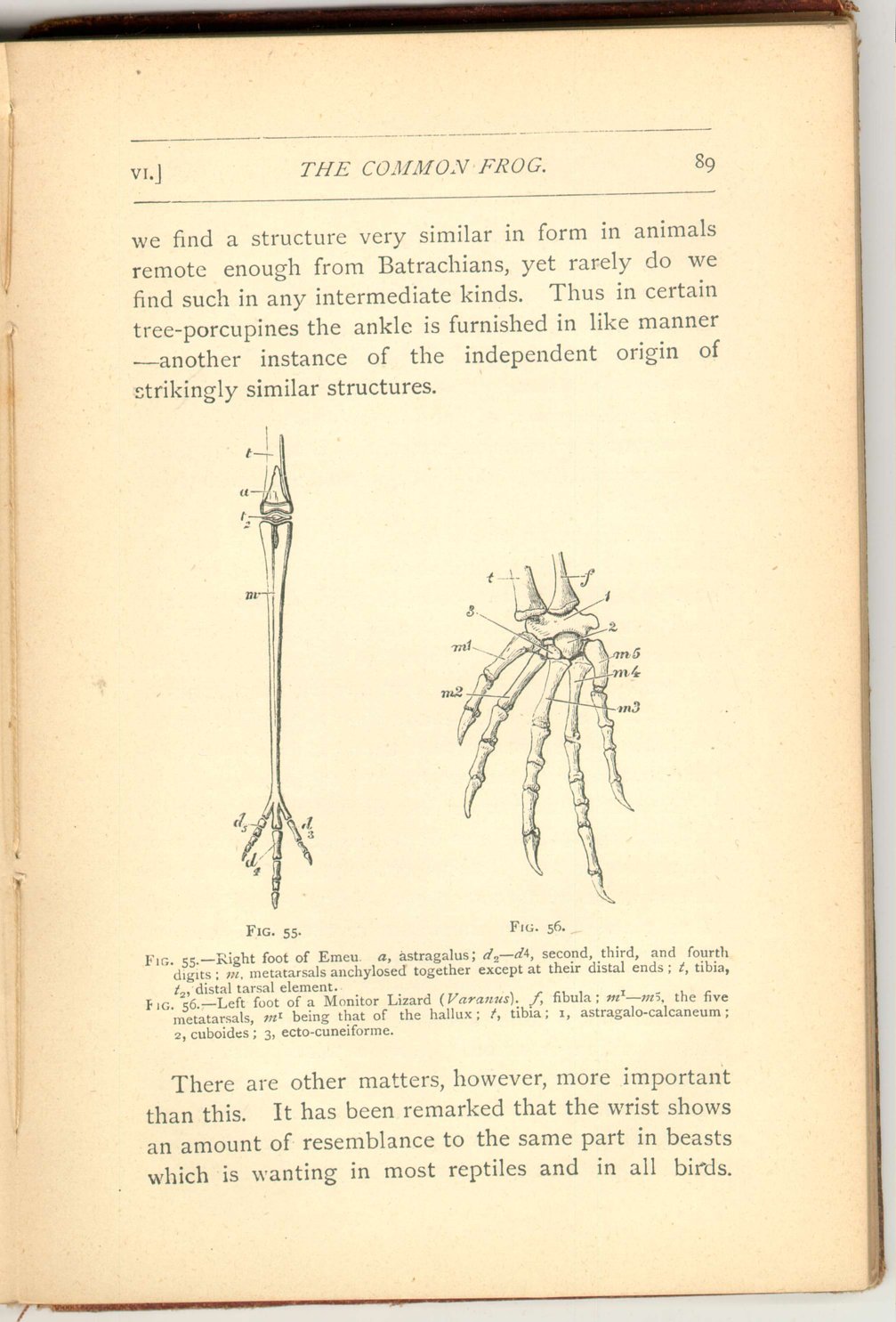
In the first place we meet in the frog with certain extra ossicles in the inner side of the foot, which present the appearance of a broad rudiment of an extra digit on the inner side of the great toe. Now
 [Description:
Image of page 89.
]
[Description:
Image of page 89.
]
There are other matters, however, more important than this. It has been remarked that the wrist shows an amount of resemblance to the same part in beasts which is wanting in most reptiles and in all birds.
 [Description:
Image of page 90.
]
[Description:
Image of page 90.
]
In all beasts, as in man, the motion of the leg on the foot takes place by means of a joint between the shin-bone of the leg and the small bones of the ankle; and though in some beasts (as in the orang) there is considerable power of motion between the first and the second row of ankle-bones, this is small compared with the mobility of the foot and ankle taken together upon the leg,
In all birds, on the contrary, not only is there no motion between the ankle-bones (as a whole) and the shin-bone, but the two rows of ankle-bones actually anchylose respectively with adjacent parts—the row nearer the leg coming to form one with the shin-bone; the second row coming to form one with the bones of the foot. Thus in birds the motion of the foot on the leg takes place not between the ankle and the shin-bone, but between the two rows of ankle-bones.
The same thing to a less degree takes place in reptiles; the ankle-bones do not indeed anchylose with the shin-bone and foot respectively, but they nevertheless unite with those parts so firmly that motion takes place between the bones of the ankle and not between the whole ankle and the leg.
Now in the frog's class, e.g. in the order Urodela, we meet with a condition which is mammalian rather than reptilian or avian. Motion takes place freely between the leg and the whole tarsus. Moreover, the number and proportions of the ankle-bones themselves far more closely agree with the condition of the same parts exhibited to us by certain beasts than
 [Description:
Image of page 91.
]
[Description:
Image of page 91.
]
The frogs and toads, however, differ from the Urodela and present us with a peculiar condition of the ankle-bones, in that the two which represent the bones of the first row are so greatly elongated as to give to the limb an additional segment—as it were two "long bones" more.
We should search in vain through every other order of the Batrachian class, through every known group of birds and reptiles, both living and fossil, to find any
 [Description:
Image of page 92.
]
[Description:
Image of page 92.
]
Amongst the very peculiar beasts which inhabit the island of Madagascar, there are certain small creatures,
 [Description:
Image of page 93.
]
[Description:
Image of page 93.
]
FOOTNOTES: Chapter 6
|
CHAPTER VI.
THE SKELETON OF THE FROG
The Common Frog | ||