|
CHAPTER IX.
THE CIRCULATION 0F THE FROG.
The Common Frog | ||
9.
CHAPTER IX.
THE CIRCULATION 0F THE FROG.
 [Description:
Image of page 128.
]
[Description:
Image of page 128.
]
Not only every animal, but
every living being, requires, in order to carry on the
functions of life, to interchange some of the gaseous
elements of its body with gases of the medium (air or
water) in which it happens to live.
Another function of extreme generality is that of conveying to all the parts and organs of the body nutritious matter for their growth or for the repair of those destructive effects which the processes of' life inevitably produce in them.
In all members of the highest sub-kingdom of animals (i.e. in all Vertebrata) these processes of gaseous interchange and nutrition are effected by means of closed vessels, along which the stream of nutritious fluid (the blood) is continually carried in a definite and constant course. During some or other part of that course the blood becomes exposed to conditions specially favourable to the gaseous interchange, the blood parting with carbonic acid gas and obtaining in its place an increased supply of oxygen.
 [Description:
Image of page 129.
]
[Description:
Image of page 129.
]
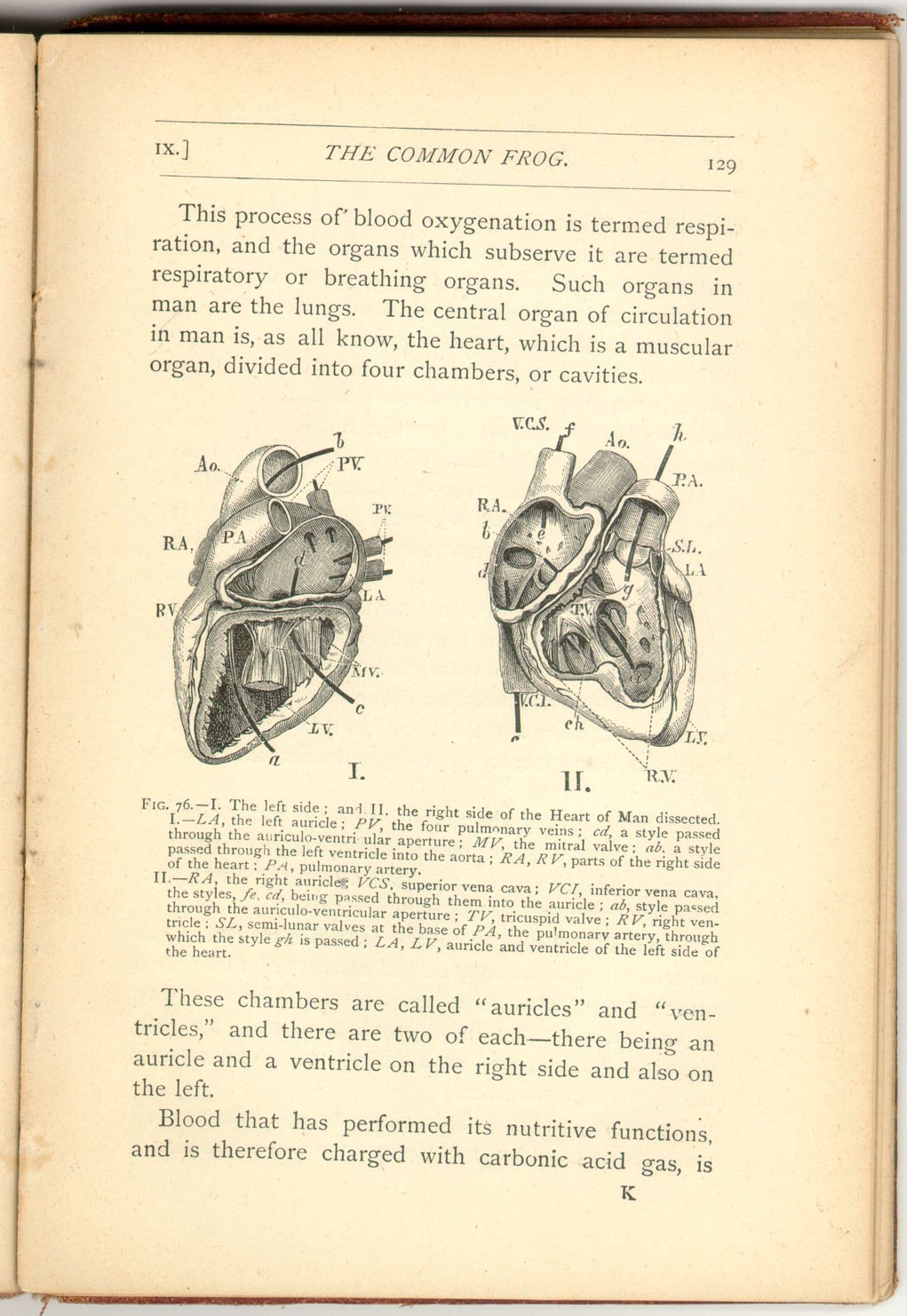
This process of blood oxygenation is termed respiration, and the organs which subserve it are termed respiratory or breathing organs. Such organs in man are the lungs. The central organ of circulation in man is, as all know, the heart, which is a muscular organ, divided into four chambers, or cavities.
Fig. 76.—I. The left side; and II. the right side of the Heart of Man dissected.
I. —LA, the left auricle; PV, the four pulmonary veins; cd, a style passed through the auriculo-ventricular aperture; MV, the mitral valve; ab, a style passed through the left ventricle into the aorta; RA, RV, parts of the right side of the heart: PA, pulmonary artery.
II.—RA, the right auricle; VCS, superior vena cava; VCI, inferior vena cava, the styles fe, cd, being passed through them into the auricle; ab, style passed through the auriculo-ventricular aperture; TV, tricuspid valve; RV, right ventricle; SL, semi-lunar valves at the base of PA, the pulmonary artery, through which the style gh is passed; LA, LV, auricle and ventricle of the left side of the heart.
These chambers are called "auricles" and "ventricles," and there are two of each—there being an auricle and a ventricle on the right side and also on the left.
Blood that has performed its nutritive functions, and is therefore charged with carbonic acid gas, is
 [Description:
Image of page 130.
]
[Description:
Image of page 130.
]
The oxygenated, or arterial blood, is returned from the lungs to the left auricle, and hence it is directly transmitted to the left ventricle, whence it is driven through the great artery (the aorta) into other arteries, and so distributed all over the body. The aorta passes downwards in front of the backbone, when it is called the descending aorta. Before turning downwards, however, it gives off great arteries to the arms and head, the carotid arteries carrying blood to the latter.
Now it is very important that the blood should not proceed in a direction the reverse of that indicated, and to prevent such misdirection, or regurgitation, special valves are placed at different openings; these valves freely allowing the blood to flow in the proper direction, but instantly opposing an effectual obstacle to a contrary flux.
The openings of the auricles into the ventricles are guarded by valves, as also is the opening of the left ventricle into the aorta, and that of the right ventricle into the artery going to the lungs.
The valve which guards the entrance into the right ventricle is called tricuspid, and consists of three flaps attached by delicate tendinous cords in such a way as to hinder the tending backwards of the flaps into the right auricle, and so allowing the blood to flow back into that chamber.
The valve which guards the entrance into the left ventricle is called mitral (from a fancied resemblance
 [Description:
Image of page 131.
]
[Description:
Image of page 131.
]
In man the whole of the blood is sent to the lungs for purification during each circuit of this most important fluid, and every organ is supplied with oxygenated blood.
If in any animals the process of purification is incomplete, it is manifestly desirable that these organs of the body, the functions of which are the most important, should be supplied with that part of the blood which is pure. This consideration eminently applies to the brain, the director and controller of the entire body.
Now all birds and beasts without exception, share with man this perfect aeration of the entire blood, the whole of the blood in the classes Mammalia and Aves being purified in the lungs before being distributed to the body.
The conditions by which the frog, at the various stages of its existence, oxygenates its blood and directs the purified stream in the most desirable manner, are curious and instructive.
It is generally known that the lower air-breathing Vertebrates (Reptiles and Batrachians ) have the heart less completely divided than in the higher classes, so that the oxygenated (or arterial) blood and the unoxygenated (or venous) blood become mixed in the single or imperfectly divided ventricle.
It might well be supposed, and in fact has generally been so, that in animals with a heart so imperfectly
 [Description:
Image of page 132.
]
[Description:
Image of page 132.
]
In fact, however, this is by no means the case, and in the frog, in spite of the reception into a single chamber of both venous blood from the body, and of arterial blood from the lungs, special mechanical arrangements effect such a definite distribution of the two sorts of blood, that the unoxygenated fluid from the body is sent to the purifying respiratory surfaces (lungs and skin), and the pure oxygenated blood alone goes to the head and to the brain.
For the detection of this beautiful mechanism we are indebted to the careful investigations of Ernst Brücke. [21]
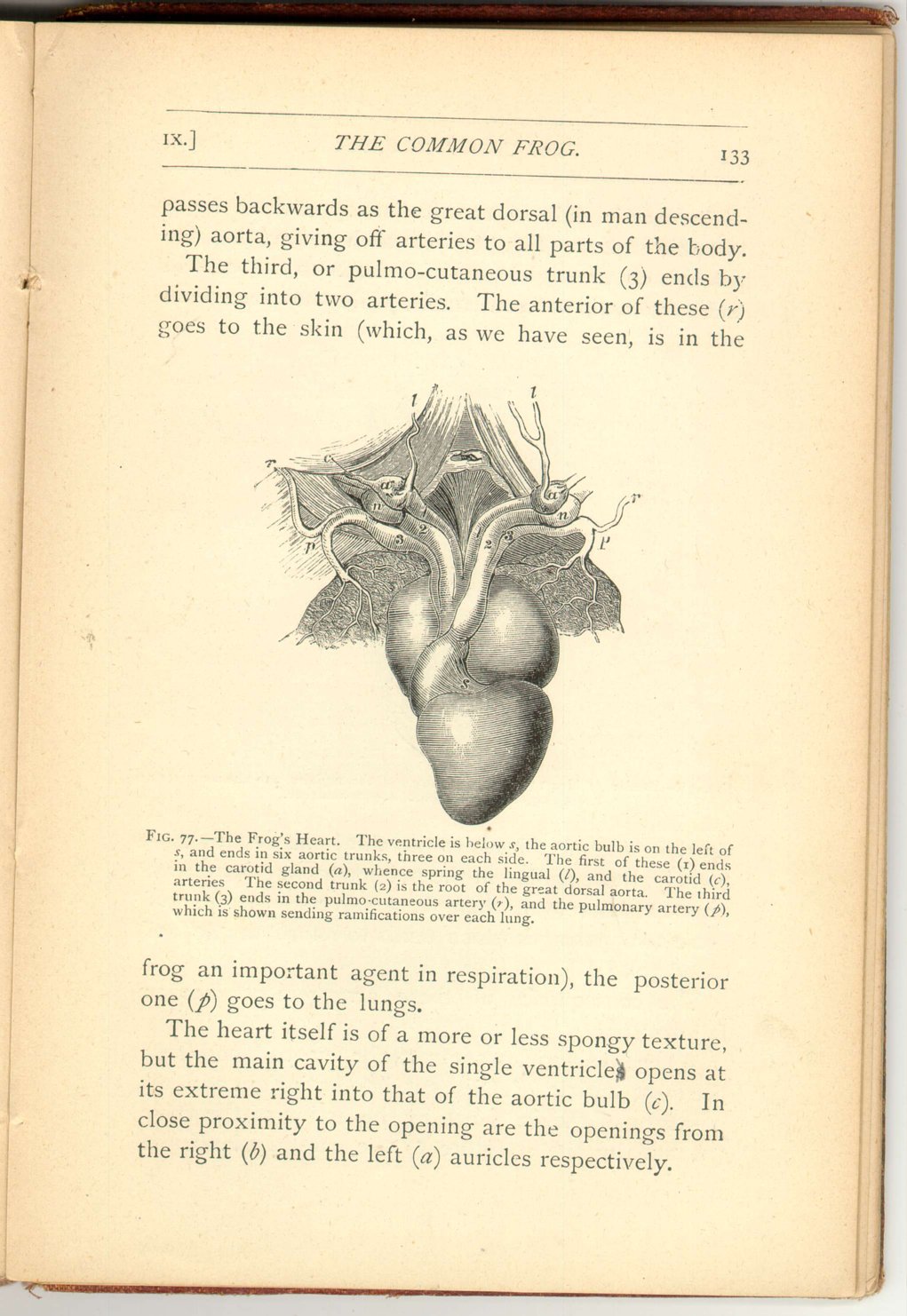
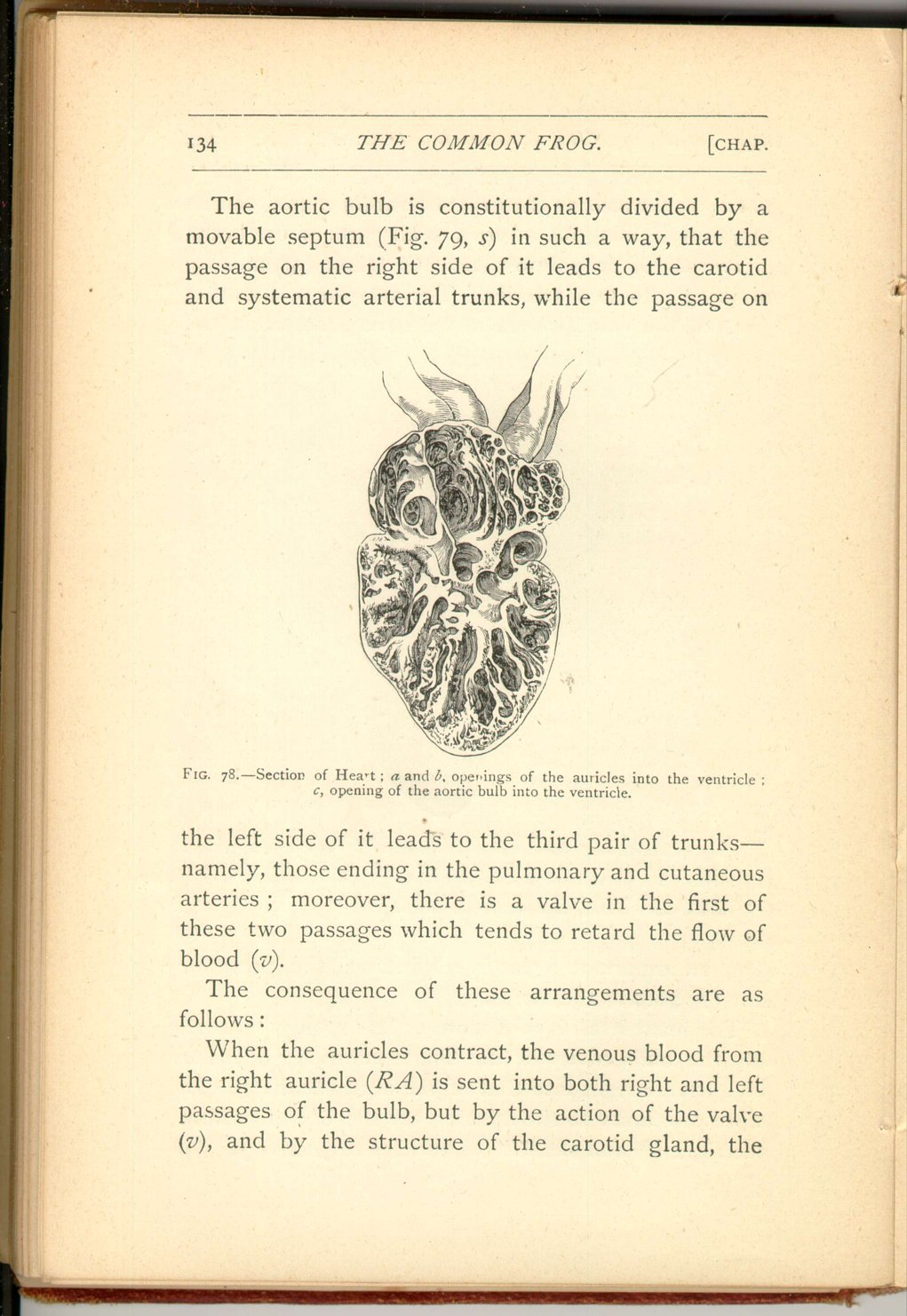
The heart of the frog consists of a right and left auricle (divided by a delicate septum), both opening into a single ventricle. From the latter proceeds an aortic root (bulbus aortæ) which gives rise to three arterial trunks on each side.
The first of these, or carotid trunk (1), ends in an enlargement (a) termed the carotid gland, of spongy structure, which gives rise to two arteries, one the lingual (l), the other (c) the carotid which goes to the head and brain.
The second, or systematic trunk (2) meets its fellow of the opposite side beneath the spine, and thence
 [Description:
Image of page 133.
]
[Description:
Image of page 133.
]
The third, or pulmo-cutaneous trunk (3) ends by dividing into two arteries. The anterior of these (r) goes to the skin (which, as we have seen, is in the
The heart itself is of a more or less spongy texture, but the main cavity of the single ventricles opens at its extreme right into that of the aortic bulb (c). In close proximity to the opening are the openings from the right (b) and the left (a) auricles respectively.
 [Description:
Image of page 134.
]
[Description:
Image of page 134.
]
The consequence of these arrangements are as follows:
When the auricles contract, the venous blood from the right auricle (RA) is sent into both right and left passages of the bulb, but by the action of the valve (v), and by the structure of the carotid gland, the
 [Description:
Image of page 135.
]
[Description:
Image of page 135.
]
As the lungs get gorged with blood, the resistance on the two sides of the septum of the bulb becomes at first equalised and soon becomes the greater on the left side; then the septum is forced over to the left, and the blood, now mixed with pure blood, flowing in from the left auricle, flows freely along the systematic arteries (2 and 2). The obstruction of the carotid glands (c gld) being the greatest and the last to be overcome, the carotid and lingual arteries (c and l) receive the very last of the blood—that, namely, which coming from the left auricle is purely arterial—
 [Description:
Image of page 136.
]
[Description:
Image of page 136.
]
So much for the circulation of the frog in its adult condition. Its larval, or tadpole stage, presents us with a series of changes which, though more familiar, are not less wonderful.
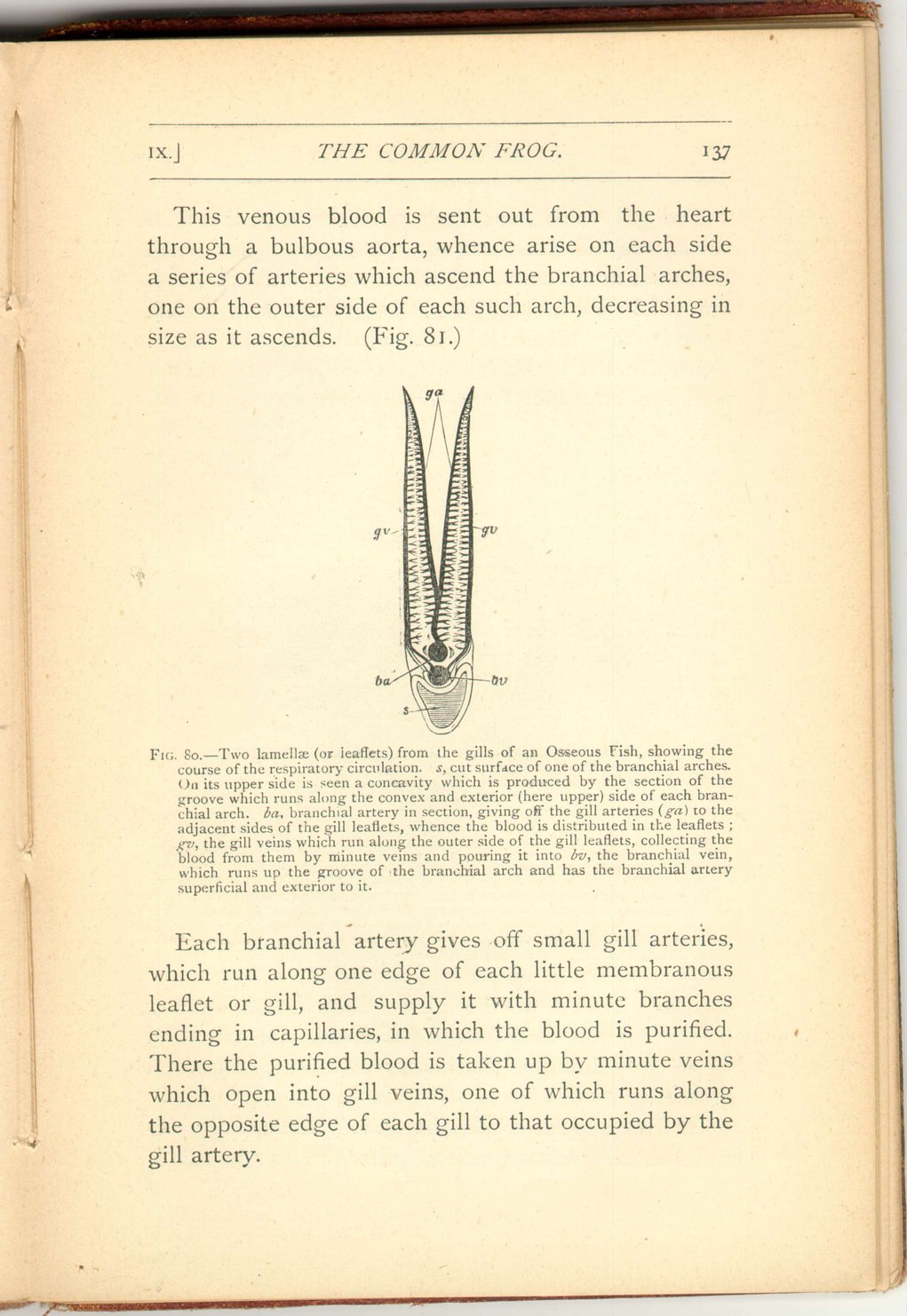
In the first place, however—it may be well to describe shortly the condition of the circulation in fishes, where the purification of the blood is effected, not by means of the exposure of the blood to the action of air taken into respiratory cavities of the body, but by its subjection in little places of membrane, the gills, to the influence of air mechanically mixed up with and dissolved in the water in which those gills are bathed.
In fishes, moreover, unlike all air-breathing animals none of the oxygenated blood is returned to the heart for propulsion, but is collected directly into the great dorsal aorta, whence it is distributed to the whole body, only being returned to the heart after such distribution, so that venous blood alone enters that organ.
 [Description:
Image of page 137.
]
[Description:
Image of page 137.
]
 [Description:
Image of page 138.
]
[Description:
Image of page 138.
]
The gill veins pour their contents into branchial veins, one of which ascends the outer side of each branchial arch, increasing in size as it ascends. The branchial veins open into the great dorsal aorta, whence the blood is distributed over the body. Generally the branchial arteries are only connected with the branchial veins by the intervention of the
In the tadpole, while the gills remain fully developed, a condition exists quite similar to that of
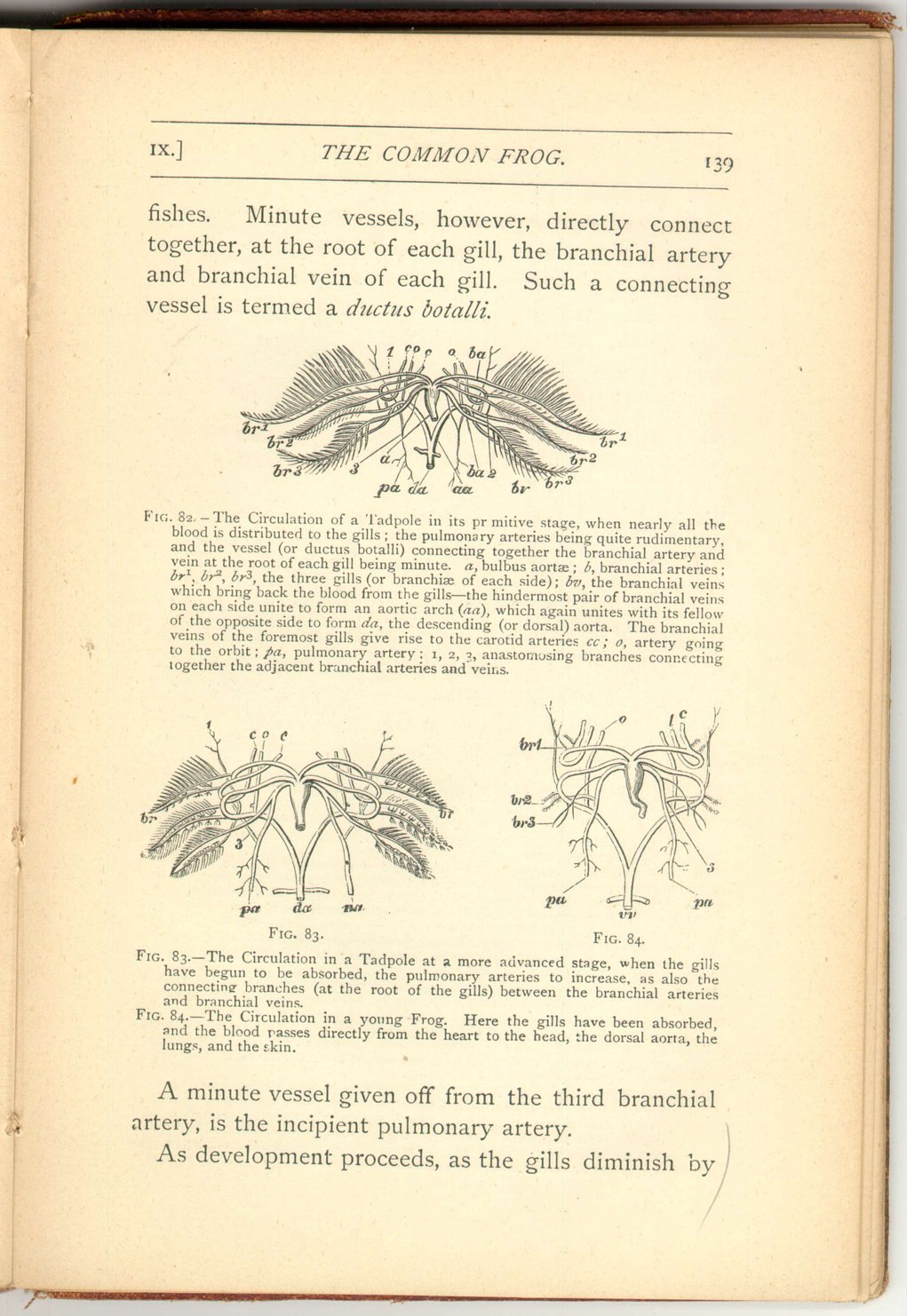 [Description:
Image of page 139.
]
[Description:
Image of page 139.
]
A minute vessel given off from the third branchial artery, is the incipient pulmonary artery.
As development proceeds, as the gills diminish by
 [Description:
Image of page 140.
]
[Description:
Image of page 140.
]
 [Description:
Image of page 141.
]
[Description:
Image of page 141.
]
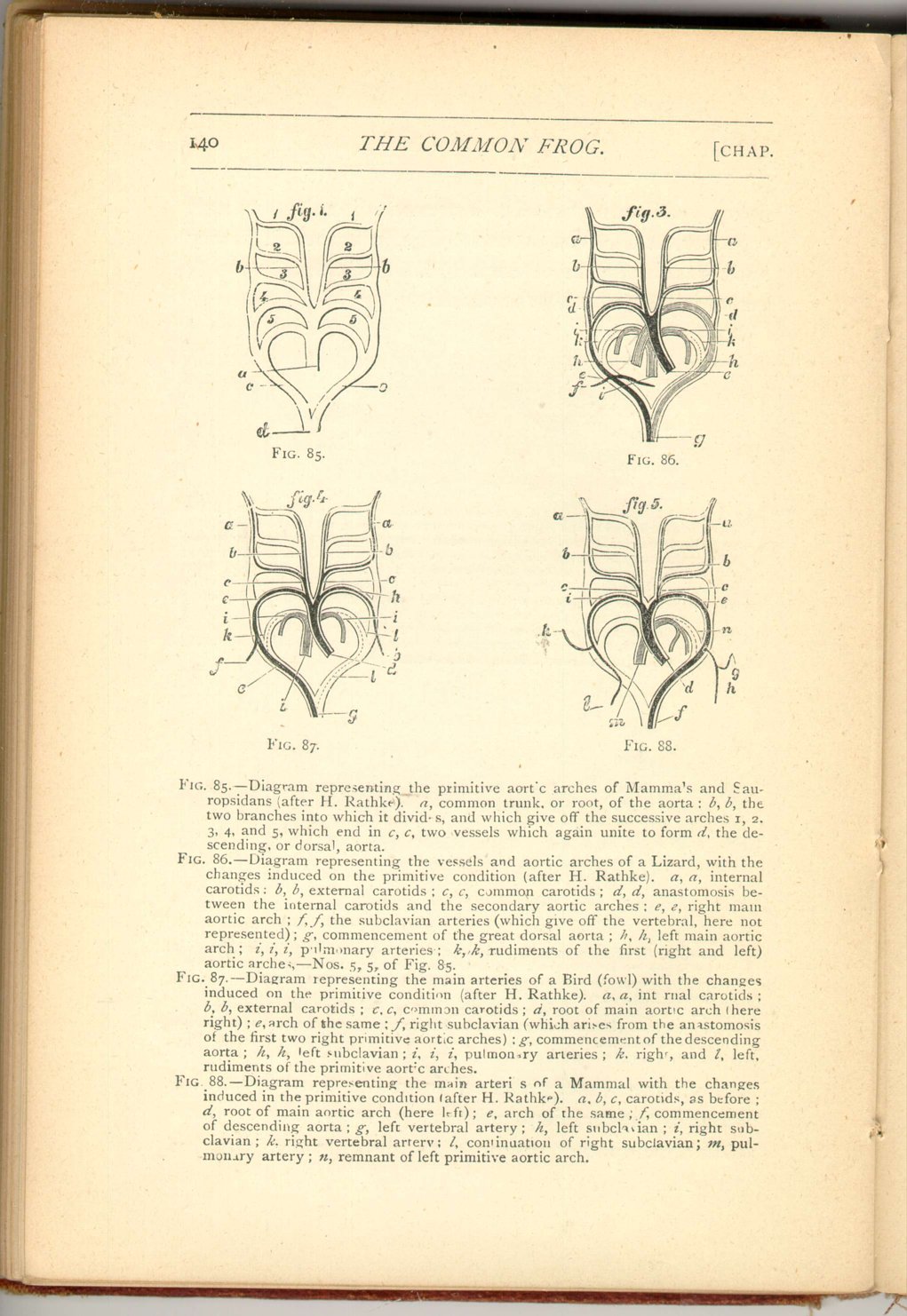
We have, then, in the life-history of the frog, a complete transition from the condition of the fish to that of a true air-breathing vertebrate, as regards its circulation. The various conditions herein referred to have, however, an important bearing on the question of the first origin of such structure.
All higher animals, even the very highest, have the great arteries when they first appear, arranged substantially as in fishes.
From the common aortic bulb five vessels ascend each side of the neck, and more or fewer of these arteries abort in different classes, the permanent adult condition being arrived at by this circuitous route.
This argument has commonly been adduced as an argument in favour of the descent of air-breathing animals from more ancient gill-bearing forms, and it is not without weight.
Nevertheless, it must be borne in mind that the primitive condition, in fishes is that of direct continuity between the branchial arteries and veins such as we have seen exists permanently in Lepidosiren. It is only as development proceeds that each primitive continuous arch becomes broken up into an artery and a vein connected by a network of capillaries.
Now we can understand the series of unbroken arches in higher animals as the relics of ancestral vessels which divided for gill-circulation and were therefore once of extreme functional importance and
 [Description:
Image of page 142.
]
[Description:
Image of page 142.
]
The frog when adult has, besides its skin, no breathing organs but the lungs. As has been said before, other members of the frog's class retain gills and aquatic respiration during the whole of life, as for example Menobranchus.
Every one, kind however, whether provided permanently with gills or not, develops lungs, and it might easily be imagined that similarly every gilled-creature which has lungs is also a Batrachian.
This, however, would be a mistake.
The Mud-fish or Lepidosiren, already referred to more than once, is furnished with both gills and lungs throughout the whole of life. On this account it has been reckoned by some naturalists to be a fish and not a Batrachian. Its fish-nature, however, has now been thoroughly established, and thus the probability of the existence of lungs within the class of fishes is also established.
But what is a lung?
A lung is a sac-like structure capable of being distended with air, supplied with venous blood direct from the heart, and sending arterial blood directly to it. Generally the whole of the blood from the lungs goes back to the heart directly, but in one Batrachian—the celebrated Proteus—a portion of the blood from the lungs finds its way not into the heart but into vessels of the general circulation. When there is an air-sac which does not both receive blood directly from and return it directly to the heart—i.e. when
 [Description:
Image of page 143.
]
[Description:
Image of page 143.
]
The transition, however, from a lung to a swim-bladder is a graduated one. We have just seen that in Proteus, though blood is returned from the lungs direct to the heart, yet that not all the blood is so returned. On the other hand in another animal, Ceratodus, though blood is not brought to its air-sac directly (which is therefore a swim-bladder and not a lung), yet for all that blood is sent from it direct to the heart.
Ceratodus (or as it is locally called "flat-head") is a fish of Queensland, closely allied to Lepidosiren , and is a very noteworthy animal apart from and in addition to its peculiarly transitional structure as regards its air-sac.
It is, indeed, the last of an ancient race, a species of the same genus (known almost exclusively by its teeth), being found fossil in strata of oolitic and triassic date. It was discovered by the Hon. W. Foster, M.C.A. Mr. Gerard Krefft, F.L.S., Curator and Secretary of the Sydney Museum, first described and figured the animal in 1870 [22] and at once correctly referred it to the genus Ceratodus, which up to that time was supposed to be entirely extinct. Its further determination was effected by
 [Description:
Image of page 144.
]
[Description:
Image of page 144.
]
Ceratodus and Lepidosiren together afford the most remarkable evidence of the persistence of the same type of structure in the Vertebrate sub-kingdom. The group to which they both belong reaches back into the very earliest epoch, which has yet afforded us any evidence whatever of the existence of fishes; while the genus Ceratodus seems to have persisted unchanged from the period of the deposition \ of the triassic strata.
The Excretory 0rgans.
As has been said, it is a necessary action for every living being, whether plant or animal (in order that it may continue to live) to eliminate certain substances, the most noteworthy being carbonic acid, which is set free by the process of respiration.
The active processes of life, however, necessarily wear out, by their activity, parts containing the gas nitrogen, which enters into the composition of every animal, and especially into every part exhibiting much vital activity.
 [Description:
Image of page 145.
]
[Description:
Image of page 145.
]
The elimination of nitrogenous excreta becomes, then, a very important process, and, indeed, in ourselves death soon ensues when its elimination is prevented by injury or disease. That part of the body which is especially devoted to the excretion of the nitrogenous waste of the tissues is the renal or urinary organs—the two kidneys. Richly supplied with blood, the kidneys have the power of straining off from the blood (i.e. secreting) nitrogenous substances, which are also in part eliminated by the skin in perspiration.
The material so strained off or secreted, is conveyed by two long tubes, the ureters, which pass down, one from each kidney, to the bladder or reservoir of the nitrogenous excretion.
In very many animals, as e.g. in birds, there is no bladder, but the ureters terminate in a chamber, the cloaca, which also receives the termination of those canals (the oviducts) down which the eggs pass in order to be "laid."
In no adult bird, however, nor indeed in any adult reptile, is there any closer connection between these two sets of canals—the ureters and oviducts—which terminate independently in the cloaca. In all such animals, however, and in beasts also, at an early stage of existence we do find a certain connection between renal ducts and the oviducts or their analogues. This coincidence is owing to the fact that in such higher animals the urinary gland which ultimately exists in the adult—namely, the kidney—is not that which primitively exists, but is a subsequent formation. The primitive renal duct is not the ureter. The primitive
 [Description:
Image of page 146.
]
[Description:
Image of page 146.
]
In fishes the primitive renal (or urinary) organ persists throughout life. It answers rather to the temporary Wolffian body than to the true kidney of higher animals. It is also a fact that in some fishes (e.g. Ceratodus and Lepidosiren) a connection does exist between the renal ducts and the oviducts, anterior to the termination of either.
Now in the frog (and in its class-fellows) this highly important excretory organ presents a noteworthy condition. In the adult frogs and toads the renal or urinary gland pours its secretion by minute canals into the renal duct. This duct, however, does not open into the cloaca, as does the ureter of birds and reptiles, but into the oviduct, the oviduct and renal duct of each side thus opening into the cloaca by a single and common aperture. An analogous condition exists in the male frog and toad, and in most members of their order.
In the genera Bombinator and Diseoglossus, however, as also in the male Urodela, a still more intimate union exists between the renal organs and those devoted to functions complementary to oviposition. The
 [Description:
Image of page 147.
]
[Description:
Image of page 147.
]
The interest of these conditions is twofold. In the first place, their existence is a point of affinity with fishes, since we find in those belonging to the order to which Lepidosiren and Ceratodus pertain, a similar connection between the same two sets of organs.
It is interesting, in the second place, because, though nothing of the kind exists in adult animals of a higher class than Batrachians, yet, as already stated, in the earlier stages in the development of such animals (even the very highest) we do find a certain analogy.
The primitive urinary gland—the Wolffian body—seems then to answer to the permanent urinary gland of Batrachians. This, together with its duct, is at first, indeed, entirely devoted to the excretory function. But, as we have seen, it ultimately more or less aborts in the adults of both sexes, and becomes an appendage, exclusively, of parts which are related directly or indirectly to oviposition.
FOOTNOTES: Chapter 9
|
CHAPTER IX.
THE CIRCULATION 0F THE FROG.
The Common Frog | ||