| The Origin and Nature of the Emotions: Miscellaneous Papers | ||
THE KINETIC SYSTEM[1]
In this paper I formulate a theory which I hope will harmonize a large number of clinical and experimental data, supply an interpretation of certain diseases, and show by what means many diverse causes produce the same end effects.
Even should the theory prove ultimately to be true, it will in the mean time doubtless be subjected to many alterations. The specialized laboratory worker will, at first, fail to see the broader clinical view, and the trained clinician may hesitate to accept the laboratory findings. Our viewpoint has been gained from a consideration of both lines of evidence on rather a large scale.
The responsibility for the kinetic theory is assumed by myself, while the responsibility for the experimental data is shared fully by my associates, Dr. J. B. Austin, Dr. F. W. Hitchings, Dr. H. G. Sloan, and Dr. M. L. Menten.[2]
Introduction
The self-preservation of man and kindred animals is effected through mechanisms which transform latent energy into kinetic energy to accomplish adaptive ends. Man appropriates from environment the energy he requires in the
The kinetic system does not directly circulate the blood, nor does it exchange oxygen and carbon dioxid; nor does it perform the functions of digestion, urinary elimination, and procreation; but though the kinetic system does not directly perform these functions, it does play indirectly an important rôle in each, just as the kinetic system itself is aided indirectly by the other systems.
The principal organs which comprise the kinetic system are the brain, the thyroid, the adrenals, the liver, and the muscles. The brain is the great central battery which drives the body; the thyroid governs the conditions favoring
Adrenalin alone, thyroid extract alone, brain activity alone, and muscular activity alone are capable of causing the body temperature to rise above the normal. The functional activity of no other gland of the body alone, and the secretion of no other gland alone, can cause a comparable rise in body temperature—that is, neither increased functional activity nor any active principle derived from the kidney, the liver, the stomach, the pancreas, the hypophysis, the parathyroids, the spleen, the intestines, the thymus, the lymphatic glands, or the bones can, per se, cause a rise in the general body temperature comparable to the rise that may be caused by the activity of the brain or the muscles, or by the injection of adrenalin or thyroid extract. Then, too, when the brain, the thyroid, the adrenals, the liver, or the muscles are eliminated, the power of the body to convert latent into kinetic energy is impaired or lost. I shall offer evidence tending to show that an excess of either internal or external environmental stimuli may modify one or more organs of the kinetic system, and that this modification may cause certain diseases. For example, alterations in the efficiency of the cerebral link may yield neurasthenia, mania, dementia; of the thyroid link, Graves' disease, myxedema; of the adrenal link, Addison's disease, cardiovascular disease.
This introduction may serve to give the line of our argument. We shall now consider briefly certain salient facts which relate to the conversion of latent into kinetic energy
The amount of latent energy which may be converted into kinetic energy for adaptive ends varies in different species, in individuals of the same species, in the same individual in different seasons; in the life cycle of growth, reproduction and decay; in the waking and sleeping hours; in disease and in activity. We shall here consider briefly the reasons for some of those variations and the mechanisms which make them possible.
Biologic Consideration of the Adaptive Variation in Amounts of Energy Stored in Various Animals
Energy is appropriated from the physical forces of nature that constitute the environment. This energy is stored in the body in quantities in excess of the needs of the moment. In some animals this excess storage is greater than in other animals. Those animals whose self-preservation is dependent on purely mechanical or chemical means of defense—such animals as crustaceans, porcupines, skunks or cobras—have a relatively small amount of convertible (adaptive) energy stored in their bodies. On the contrary, the more an animal is dependent on its muscular activity for self-preservation, the more surplus available (adaptive) energy there is stored in its body. It may be true that all animals have approximately an equal amount per kilo of chemical energy—but certainly they have not an equal amount stored in a form which is available for immediate conversion for adaptive ends.
Adaptive Variation in the Rate of Energy Discharge
What chance for survival would a skunk have without odor; a cobra without venom; a turtle without carapace; or a porcupine shorn of its barbs, in an environment of powerful and hostile carnivora? And yet in such an hostile environment many unprotected animals survive by their muscular power of flight alone. It is evident that the provision for the storage of "adaptive" energy is not the only evolved characteristic which relates to the energy of the body. The more the self-preservation of the animal depends on motor activity, the greater is the range of variation in the rate of discharge of energy. The rate of energy discharge is especially high in animals evolved along the line of hunter and hunted, such as the carnivora and the herbivora of the great plains.
Influences That Cause Variation in the Rate of Output of Energy in the Individual
Not only is there a variation in the rate of output of energy among various species of animals, but one finds also variations in the rate of output of energy among individuals of the same species. If our thesis that men and animals are mechanisms responding to environmental stimuli be correct, and further, if the speed of energy output be due to changes in the activating organs as a result of adaptive stimulation, then we should expect to find physical changes in the activating glands during the cycles of increased activation. What are the facts? We know that most animals have breeding seasons evolved as adaptations to the food supply and weather. Hence there is in most animals a mating season
We have seen that there are variations in speed in different species, and that in the same species speed varies with the season of the year and with the time of day. In addition there are variations also in the rate of discharge of energy in the various cycles of the life of the individual. The young are evolved at high speed for growth, so that as soon as possible they may attain to their own power of self-defense; they must adapt themselves to innumerable bacteria, to
The cycle of greatest kinetic energy for physiologic ends is the period of reproduction. In the female especially there is a cycle of increased activity just prior to her development into the procreative state. During this time secondary sexual characters are developed—the pelvis expands, the ovaries and the uterus grow rapidly, the mammary glands develop. Again in this period of increasing speed in the expenditure of energy we find the thyroid, the adrenals, and the hypophysis also in rapid growth. Without the normal development of the ovary, the thyroid, and the hypophysis, neither the male nor the female can develop the secondary sexual characters, nor do they develop sexual desire nor show seasonal cycles of activity, nor can they procreate. The secondary sexual characters—sexual desire, fertility—may be developed at will, for example, by feeding thyroid products from alien species to the individual deprived of the thyroid.
At the close of the child-bearing period there is a permanent diminution of the speed of energy discharge, for energy is no longer needed as it was for the self-preservation of the offspring before adolescence, and for the propagation of the species during the procreative period. Unless other factors intervene, this reduction in speed is progressive until senescent death. The diminished size of the thyroid of the aged bears testimony to the part the activating organs bear in the general decline.
We have now referred to variations in the rate of discharge of energy in different species; in individuals of the same species; in cycles in the same individual—such as the seasons of food supply, the periods of wakefulness and of sleep, the procreative period, and we have spoken of those variations caused artificially by thyroid feeding, thus far having confined our discussion to the conversion for adaptive purposes of latent into kinetic energy in muscular and in procreative action. We shall now consider the conversion of latent into kinetic energy in the production of heat,[3] and endeavor to answer the questions which arise at once: Is there one mechanism for the conversion of latent energy into heat and another mechanism for its conversion into muscular action? What is the adaptive advantage of fever in infection?
The Purpose and the Mechanism of Heat Production in Infections
Vaughan has shown that the presence in the body of any alien protein causes an increased production of heat, and that there is no difference between the production of fever by foreign proteins and by infections. Before the day of the hypodermic needle and of experimental medicine, the foreign proteins found in the body outside the alimentary tract were brought in by invading microörganisms. Such organisms interfered with and destroyed the host. The body, therefore, was forced to evolve a means of protection against these hostile organisms. The increased metabolism and fever in infection might operate as a protection in two ways—the increased fever, by interfering with bacterial growth, and the
As to the mechanism which produces fever, we postulate that it is the same mechanism as that which produces muscular activity. Muscular activity is produced by the conversion of latent energy into motion, and fever is produced largely in the muscles by the conversion of latent energy into heat. We should, therefore, find similar changes in the brain, the adrenals, the thyroid, and the liver, whatever may be the purpose of the conversion of energy—whether for running, for fighting, for the expression of emotion, or for combating infection.
We shall first present experimental and clinical evidence which tends to show what part is played by the brain in the production of both muscular and febrile action, and later we shall discuss the parts played by the adrenals, the thyroid, and the liver.
Histologic Changes in the Brain-cells in Relation to the Maintenance of Consciousness and to the Production of the Emotions, Muscular Activity, and Fever
We have studied the brain-cells in human cases of fever, and in animals after prolonged insomnia; after the injection of the toxins of gonococci, of streptococci, of staphylococci, and of colon, tetanus, diphtheria, and typhoid bacilli; and after the injection of foreign proteins, of indol and skatol, of leucin, and of peptones. We have studied the brains of animals which had been activated in varying degrees up to the point of complete exhaustion by running, by fighting, by rage and fear, by physical injury, and by the injection of strychnin (Figs. 2, 4, 5, and 37). We have studied the brains of salmon at the mouth of the Columbia River and at its headwater (Fig. 55); the brains of electric fish, the storage batteries of which had been partially discharged, and of those the batteries of which had been completely discharged; the brains of woodchucks in hibernation and after fighting; the brains of humans who had died from anemia resulting from hemorrhage, from acidosis, from eclampsia, from cancer and from other chronic diseases (Figs. 40 to 43, 56, 74, and 75). We have studied also the brains of animals after the excision of the adrenals, of the pancreas, and of the liver (Figs. 57 and 60).
In every instance the loss of vitality—that is, the loss of the normal power to convert potential into kinetic energy—was accompanied by physical changes in the brain-cells (Figs. 45 and 46). The converse was also true, that is, the brain-cells of animals with normal vital power showed no histologic changes. The changes in the brain-cells were
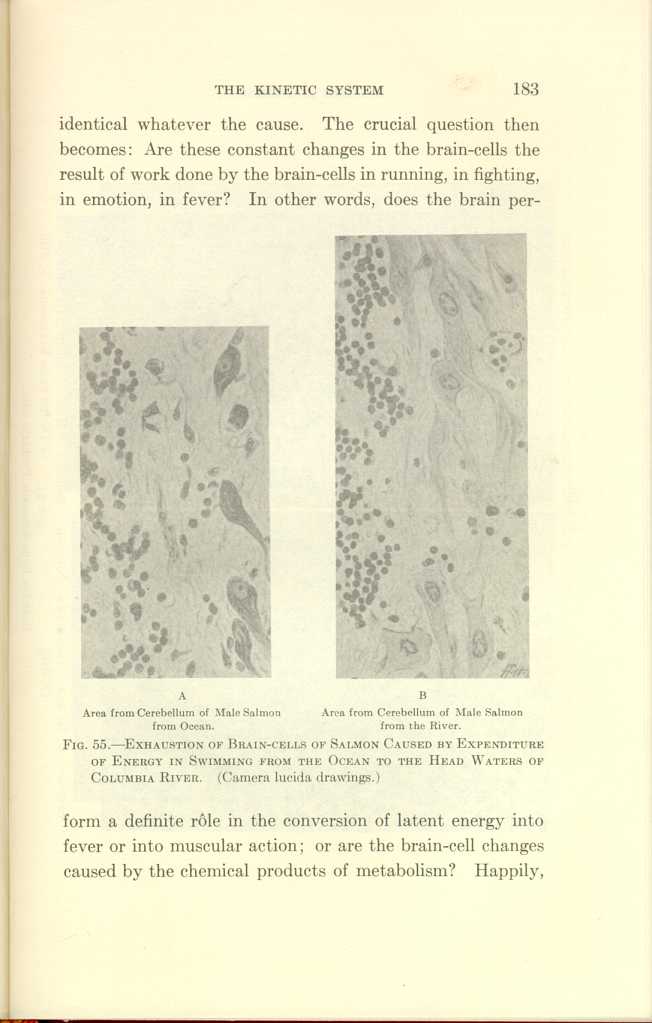
A: Area from Cerebellum of Male Salmon from Ocean.
B:Area from Cerebellum of Male Salmon from the River.
FIG. 55—EXHAUSTION OF BRAIN-CELLS OF SALMON CAUSED BY EXPENDITURE
OF ENERGY IN SWIMMING FROM THE OCEAN TO THE HEAD WATERS OF
COLUMBIA RIVER. (Camera lucida drawings.)
[Description: Black-and-white photographs showing cerebellum of salmon under
various conditions.]
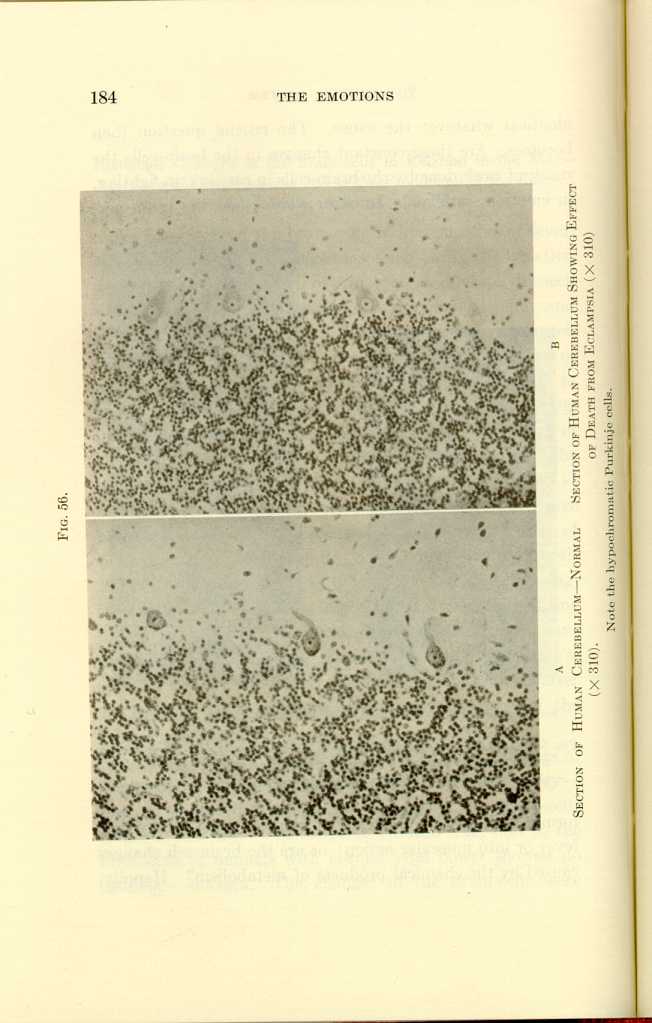
FIG. 56.
A: Section of Human Cerebellum—Normal (x310).
B: Section of Human Cerebellum Showing Effect of Death from Eclampsia
(x310).[a]
[Description: Black-and-white photographs showing human cerebellum under
various conditions.]
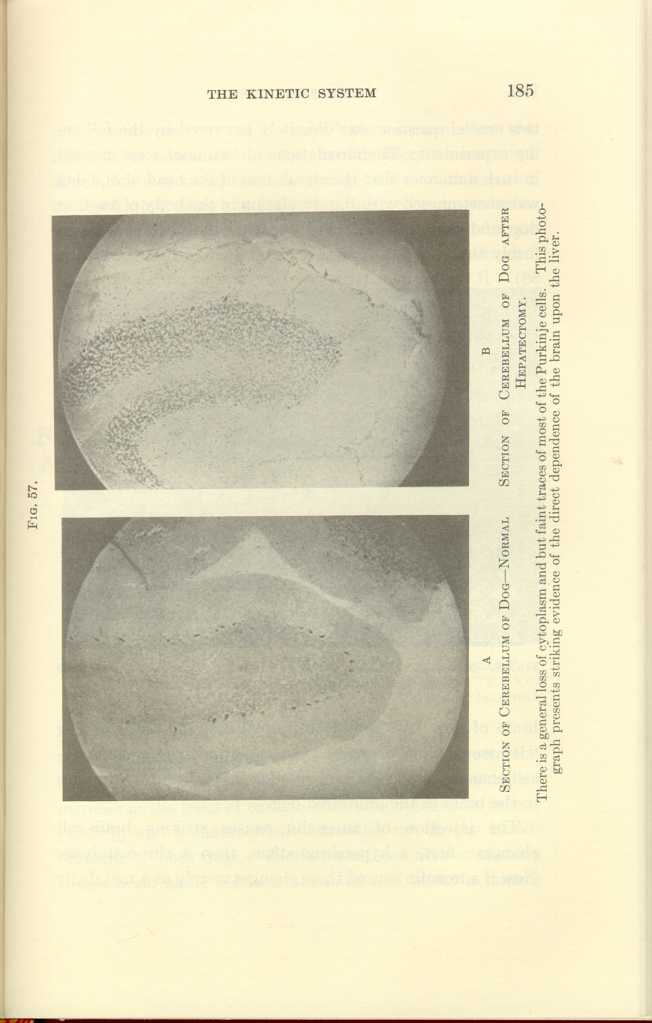
FIG. 57.
A: Section of Cerebellum of Dog—Normal
B: Section of Cerebellum of Dog after
Hepatectomy.[b]
[Description: Black-and-white photographs showing cerebellum of dog under
various conditions.]
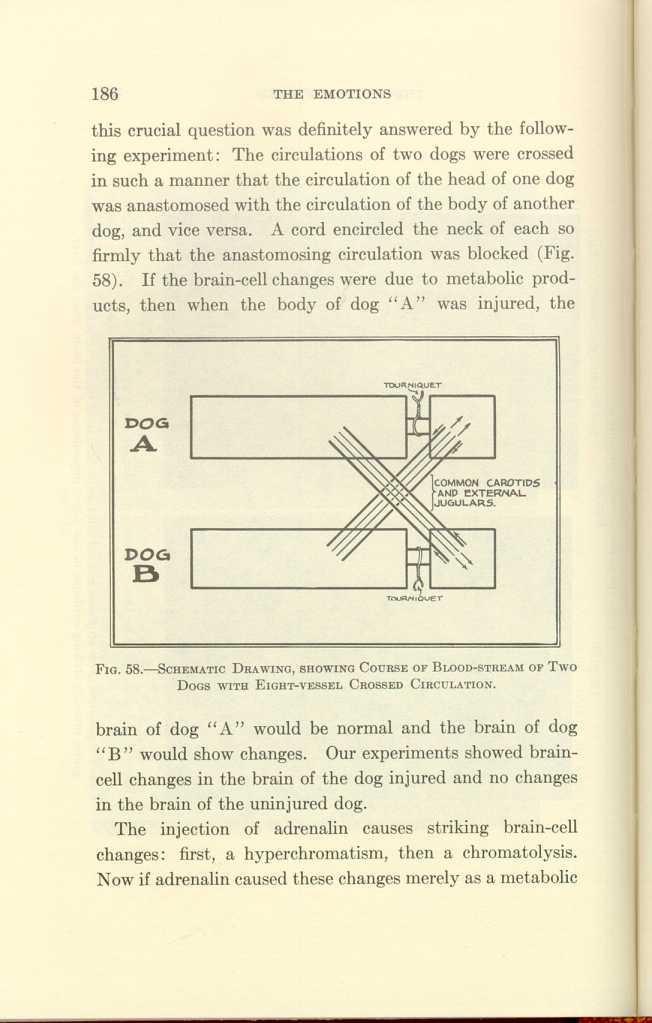
FIG. 58.—SCHEMATIC DRAWING, SHOWING COURSE OF BLOOD-STREAM OF TWO DOGS WITH EIGHT-VESSEL CROSSED CIRCULATION.
[Description: Black-and-white illustration: a diagram.]The injection of adrenalin causes striking brain-cell changes: first, a hyperchromatism, then a chromatolysis. Now if adrenalin caused these changes merely as a metabolic
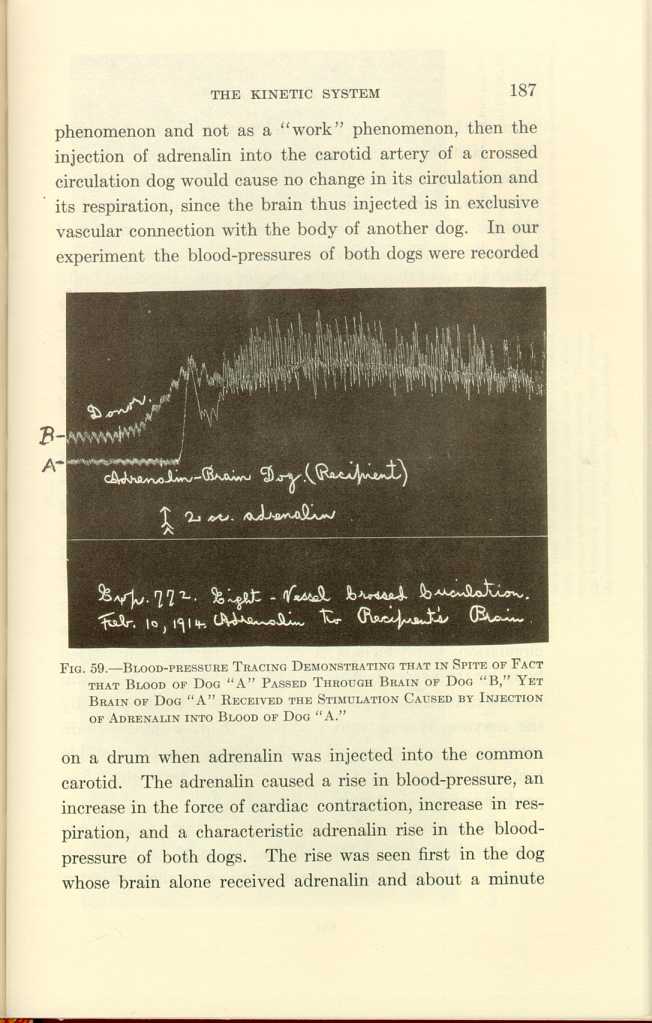
FIG. 59.—BLOOD-PRESSURE TRACING DEMONSTRATING THAT IN SPITE OF FACT THAT BLOOD OF DOG "A" PASSED THROUGH BRAIN OF DOG "B," YET BRAIN OF DOG "A" RECEIVED THE STIMULATION CAUSED BY INJECTION OF ADRENALIN INTO BLOOD OF DOG "A."
[Description: Black-and-white photograph of a chart.]1. Adrenalin alone causes hyperchromatism followed by chromatolysis, and in overdosage causes the destruction of some brain-cells.
2. When both adrenal glands are excised and no other factor is introduced, the Nissl substance progressively disappears from the brain-cells until death. This far-reaching point will be taken up later (Fig. 60).
Here our purpose is to discuss the cause of the brain-cell changes. We have seen that in crossed brain and body circulation trauma causes changes in the cells of the brain which is disconnected from the traumatized body by its circulation, but which is connected with the traumatized body by the nervous system. We have seen that adrenalin causes activation of the body connected with its brain by the nervous system, and histologic changes in the brain acted on directly by the adrenalin, but we found no notable brain-cell changes in the other brain through which the products of metabolism have circulated.
In the foregoing we find direct evidence that the products of metabolism are not the principal cause of the brain-cell changes. We shall now present evidence to show that for
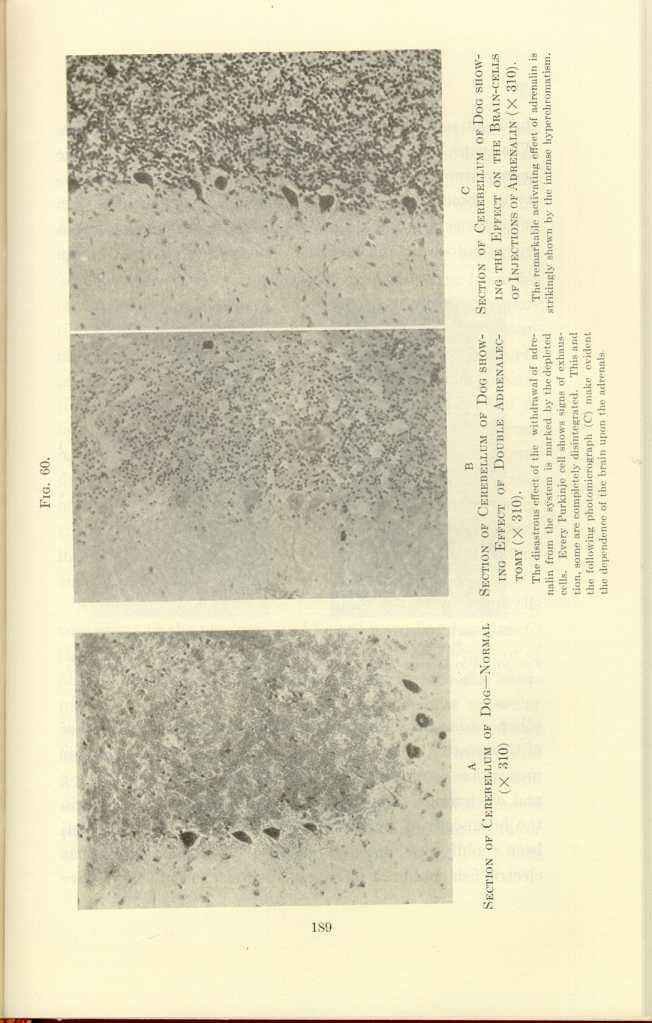
FIG. 60
A: Section of Cerebellum of Dog—Normal (x310).
B: Section of Cerebellum of Dog showing Effect of Double Adrenalectomy
(x310).[c]
C: Section of Cerebellum of Dog showing the Effect on the Brain-cells
of Injections of Adrenalin (x310).[d]
[Description: Black-and-white photographs showing cerebellum of dog under
various conditions.]
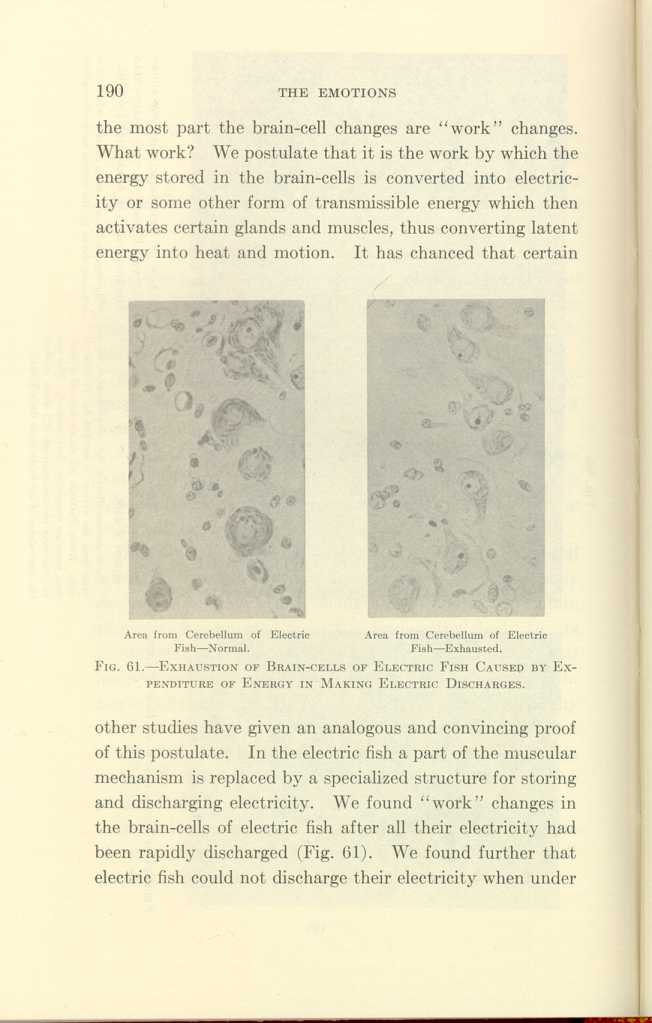
Area from Cerebellum of Electric Fish—Normal.
Area from Cerebellum of Electric Fish—Exhausted.
FIG. 61-EXHAUSTION OF BRAIN-CELLS OF ELECTRIC FISH CAUSED BY EXPENDITURE
OF ENERGY IN MAKING ELECTRIC DISCHARGES.
[Description: Black-and-white photographs showing cerebellum of fish under
normal and exhausted conditions.]
A further support to the postulate that the brain-cells contribute to the production of fever by sending impulses to the muscles is found in the effect of muscular exertion, or of other forms of motor stimulation, in the presence of a fever-producing infection. Under such circumstances muscular exertion causes additional fever, and causes also added but identical changes in the brain-cells. Thyroid extract and iodin have the same effect as muscular exertion and infection in the production of fever and the production of brain-cell changes. All this evidence is a strong argument in favor of the theory that certain constituents of the brain-cells
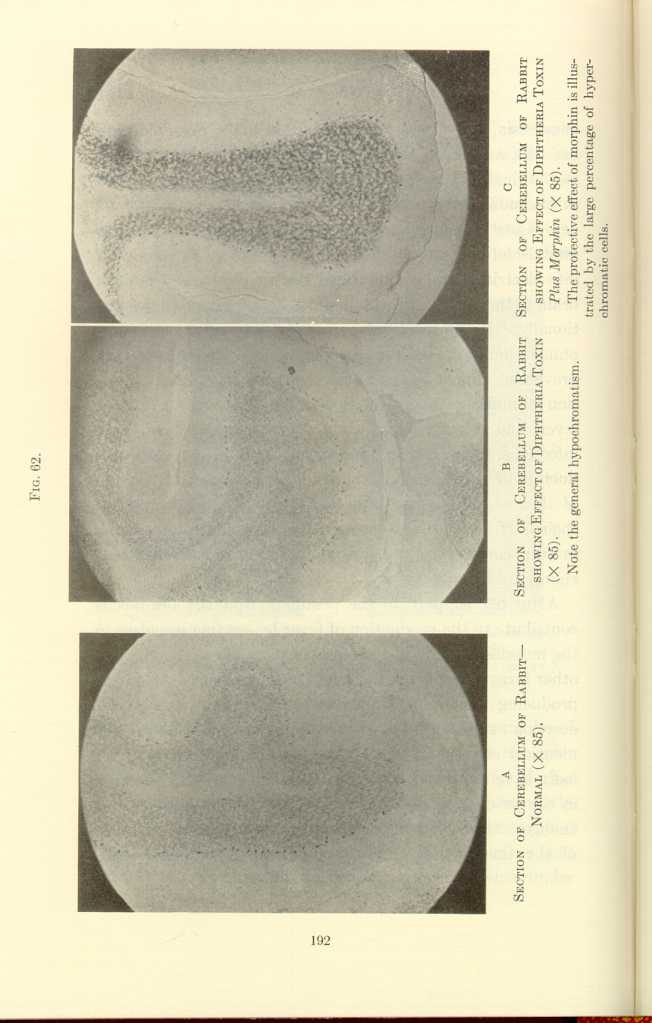
FIG. 62.
A: Section of Cerebellum of Rabbit—Normal (x85).
B: Section of Cerebellum of Rabbit showing Effect of Diptheria Toxin
(x85).[e]
C: Section of Cerebellum of Rabbit showing Effect of Diptheria Toxin
Plus Morphin (x85).[f]
[Description: Black-and-white photographs showing cerebellum of rabbit under
various conditions.]
That the stimulation of the brain-cells without gross activity of the skeletal muscles and without infection can produce heat is shown as follows:
(a) Fever is produced when animals are subjected to fear without any consequent exertion of the skeletal muscles.
(b) The temperature of the anxious friends of patients
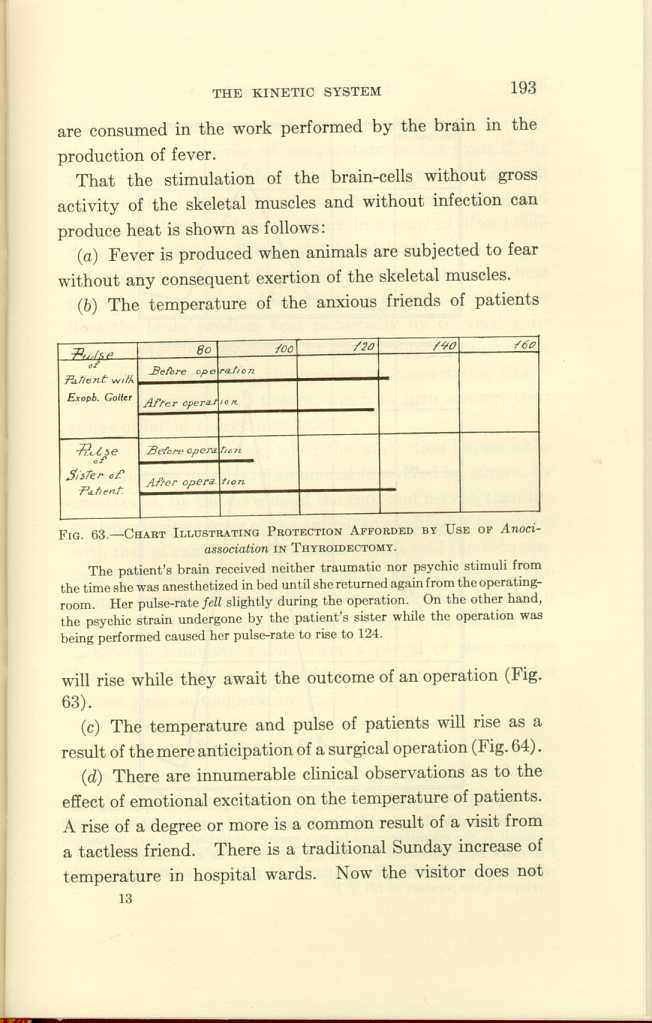
FIG. 63.-CHART ILLUSTRATING PROTECTION AFFORDED BY USE OF
Anoci-association IN THYROIDECTOMY.[g]
[Description: Black-and-white illustration: chart.]
(c) The temperature and pulse of patients will rise as a result of the mere anticipation of a surgical operation (Fig. 64).
(d) There are innumerable clinical observations as to the effect of emotional excitation on the temperature of patients. A rise of a degree or more is a common result of a visit from a tactless friend. There is a traditional Sunday increase of temperature in hospital wards. Now the visitor does not
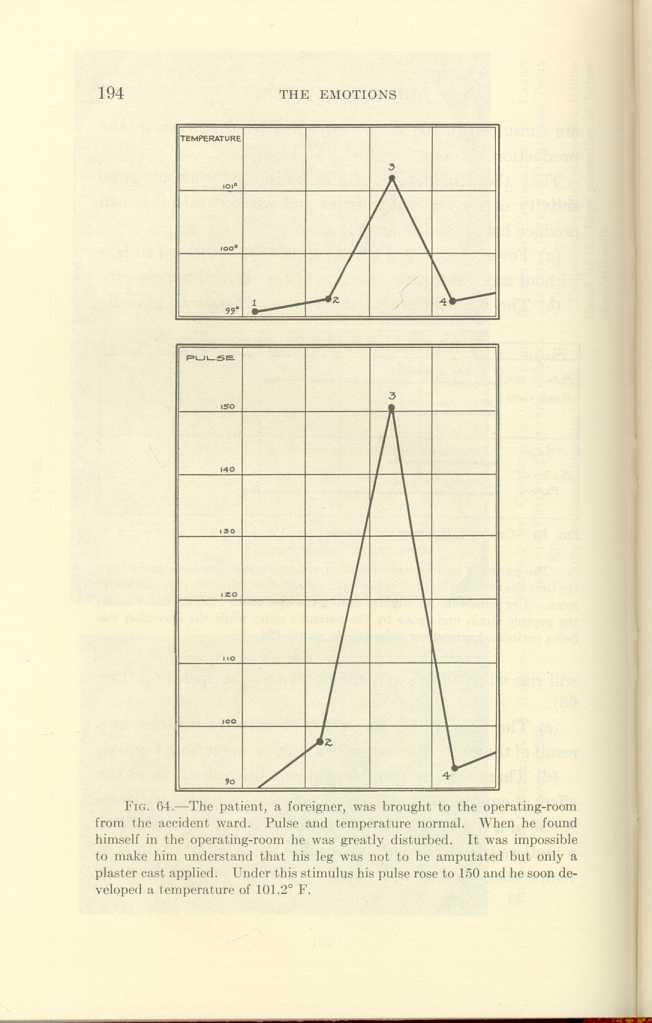
FIG. 64.[h]
[Description: Black-and-white illustration: two charts showing body temperature and pulse.]Is the contribution of the brain to the production of heat due to the conversion of latent energy directly into heat, or does the brain produce heat principally by converting its latent energy into electricity or some similar form of transmissible energy which, through nerve connections, stimulates other organs and tissues, which in turn convert their stores of latent energy into heat?
According to Starling, when the connection between the brain and the muscles of an animal is severed by curare, by anesthetics, by the division of the cord and nerves, then the heat-producing power of the animal so modified is on a level with that of cold-blooded animals. With cold the temperature falls, with heat it rises. Such an animal has no more control over the conversion of latent energy into heat than it has over the conversion of latent energy into motion.
Electric stimulation done over a period of time causes brain-cell changes, and electric stimulation of the muscles causes a rise in temperature.
Summary of Brain-cell Studies
In our crossed circulation experiments we found that neither waste products nor metabolic poisons could be considered the principal cause of the brain-cell changes. We found that in the production both of muscular action and of fever there were brain-cell changes which showed a quantitative
We shall next consider other organs of the kinetic system in their relation to muscular activity, to emotion, to consciousness, to sleep, to hibernation, and to heat production.
The Adrenals
In our extensive study of the brain in its relation to the production of energy and the consequent exhaustion caused by fear and rage; by the injection of foreign proteins, of bacterial toxins, and of strychnin; by anaphylaxis; by the injection of thyroid extract, of adrenalin, and of morphin, we found that, with the exception of morphin, each of these agents produced identical changes in the brain-cells. As we believed that the adrenals were intimately associated with the brain in its activities, we concluded that the adrenals also must have been affected by each of these agents. To prove this relation, we administered the above-mentioned stimuli to animals and studied their effects upon the adrenals by functional, histologic, and surgical methods, the functional tests being made by Cannon's method.
Functional Study of the Adrenals.—Our method of applying the Cannon test for adrenalin was as follows: (a) The blood of the animals was tested before the application of the stimulus. If this test was negative, then (b) the
We have recorded 66 clear-cut experiments on dogs, which show that after fear and rage, after anaphylaxis, after injections of indol and skatol, of leucin and creatin, of the toxins of diphtheria and colon bacilli, of streptococci and staphylococci, of foreign proteins, and of strychnin, the Cannon test for adrenalin was positive. The test was negative after trauma under anesthesia, and after intravenous injections of thyroid extract, of thyroglobin, and of the juices of various organs injected into the same animal from which the organs were taken. Placental extract gave a positive test. The test was sometimes positive after electric stimulation of the splanchnic nerves. On the other hand, if the nerve supply to the adrenals had been previously divided, or if the adrenals had been previously excised, then the Cannon test was negative after the administration of each of the foregoing adequate stimuli. Blood taken directly from the adrenal vein gave a positive result, but under deep morphinization the blood from the adrenal vein was negative, and under deep morphinization the foregoing adequate stimuli were negative.
In brief, the agencies that in our brain-cell studies were
Histologic Study of the Adrenals.—Histologic studies of the adrenals after the application of the adequate stimuli which gave positive results to the Cannon test for adrenalin are now in progress, and thus far the histologic studies corroborate the functional tests.
In hibernating woodchucks, the cells of the adrenal cortex were found to be vacuolated and shrunken. In one hundred hours of insomnia, in surgical shock, in strong fear, in exhaustion from fighting, after peptone injections, in acute infections, the adrenals undergo histologic changes characteristic of exhaustion (Figs. 66 to 67).
We have shown that brain and adrenal activity go hand in hand, that is, that the adrenal secretion activates the brain, and that the brain activates the adrenals. The fundamental question which now arises is this: Are the brain and the adrenals interdependent? A positive answer may be given to this question, for the evidence of the dependence of the brain upon the adrenals is as clear as is the evidence of the dependence of the adrenals upon the brain. (1) After excision of the adrenals, the brain-cells undergo continuous histologic and functional deterioration until death. During this time the brain progressively loses its power to respond to stimuli and there is also a progressive loss of muscular power and a diminution of body temperature. (2)
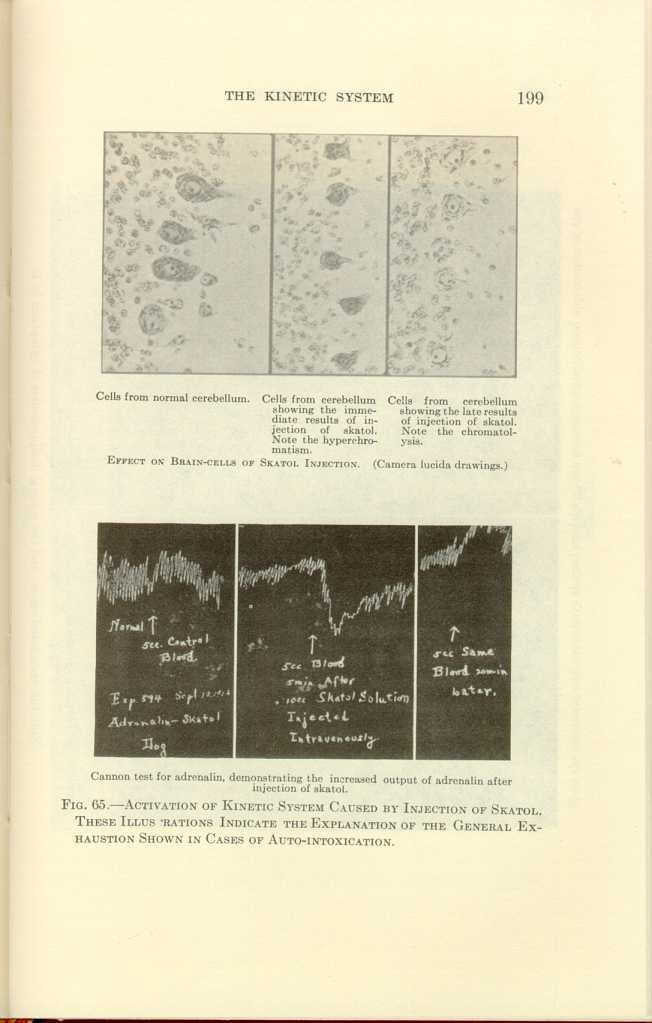
Cells from normal cerebellum.
Cells from cerebellum showing the immediate results of injection of
skatol. Note the hyperchromatis.
Cells from cerebellum showing the late results of injection of skatol.
Note the chromatolysis.
EFFECT ON BRAIN-CELLS OF SKATOL INJECTION. (Camera lucida drawings.)
[Description: Black-and-white photographs showing cerebellum under various
conditions.]
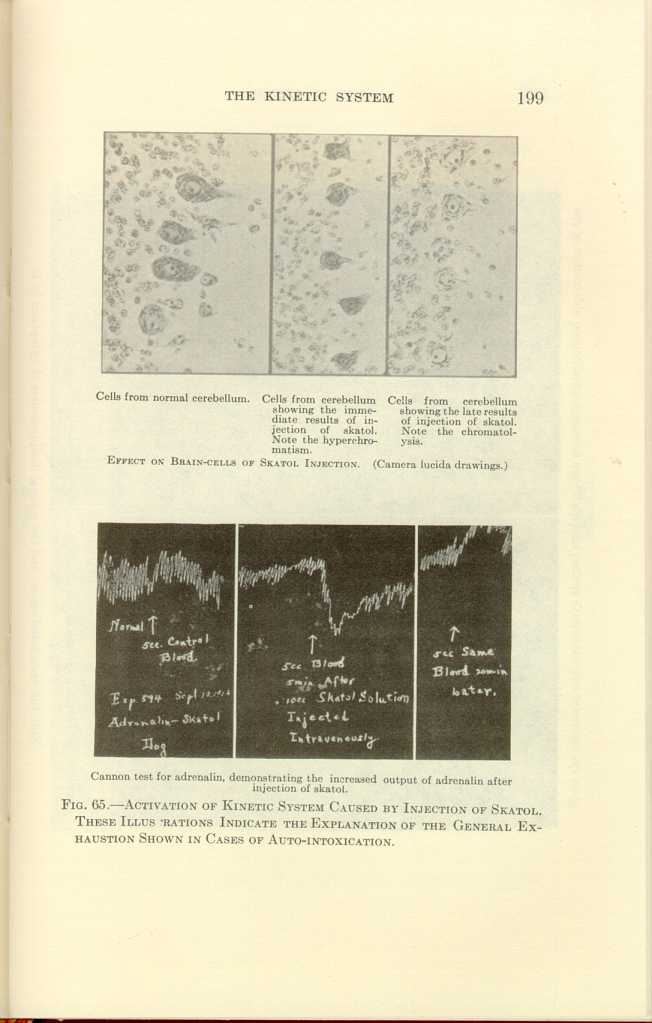
Cannon test for adrenalin, demonstrating the increased output of
adrenalin after injection of skatol.
FIG. 65.—ACTIVATION OF KINETIC SYSTEM CAUSED BY INJECTION OF SKATOL.
THESE ILLUSTRATIONS INDICATE THE EXPLANATION OF THE GENERAL EXHAUSTION
SHOWN IN CASES OF AUTO-INTOXICATION.
[Description: Black-and-white photograph of a chart.]
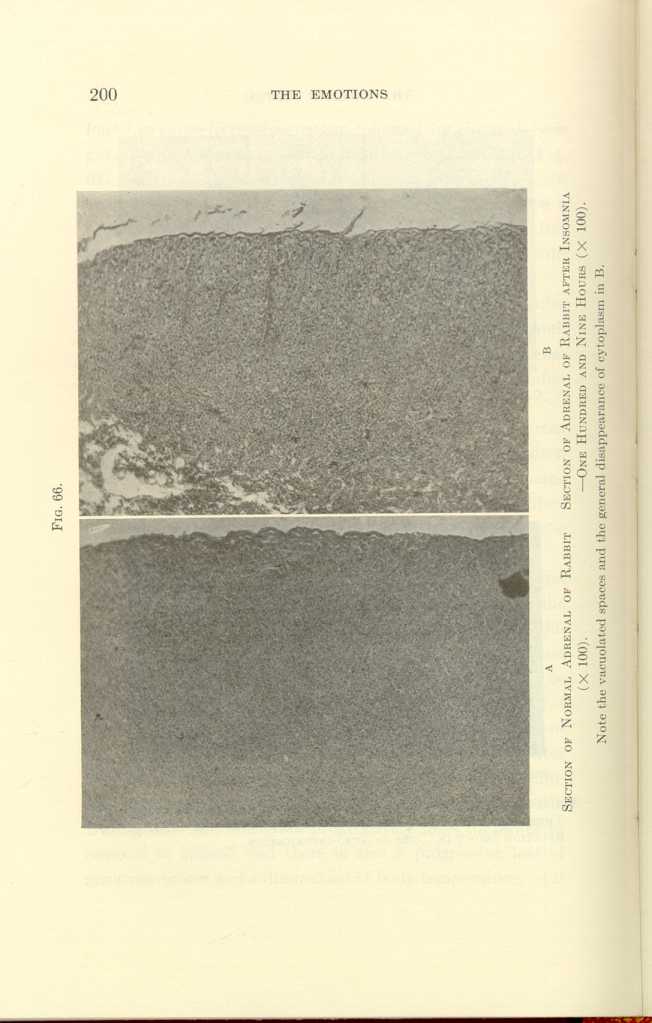
FIG. 66.
A: Section of Normal Adrenal of Rabbit (x100).
B: Section of Adrenal of Rabbit after Insomnia—One Hundred and Nine
Hours (x100).[i]
[Description: Black-and-white photographs showing adrenal of rabbit under
various conditions.]
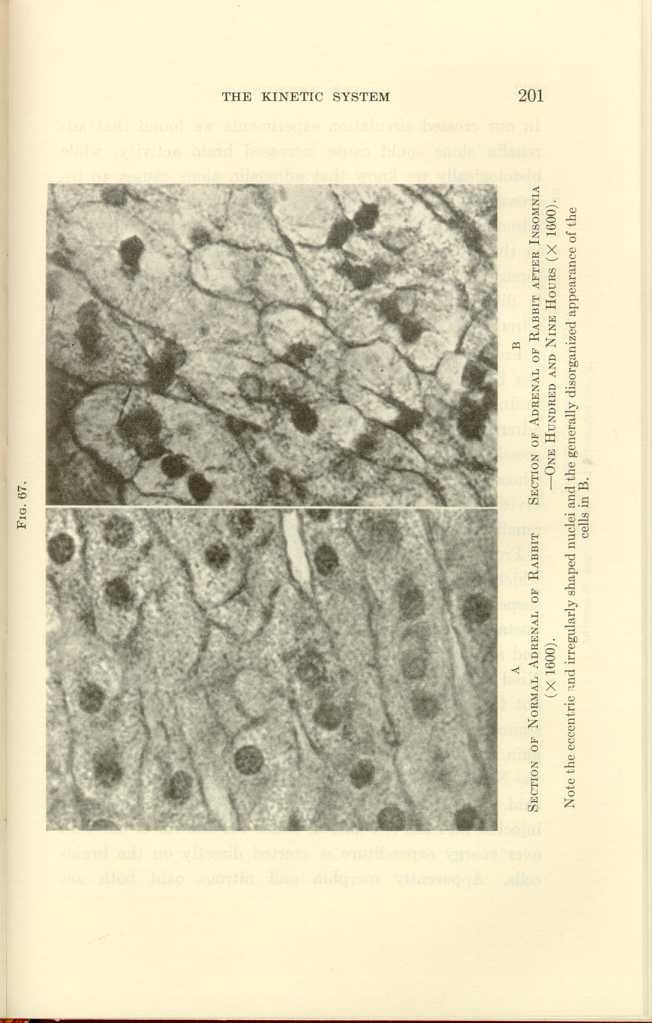
FIG. 67.
A: Section of Normal Adrenal of Rabbit (x1600).
B: Section of Adrenal of Rabbit after Insomnia—One Hundred and Nine
Hours (x1600).[j]
[Description: Black-and-white photographs showing adrenal of rabbit under
various conditions.]
From these studies we are forced to conclude not only that the brain and adrenals are interdependent, but that the brain is actually more dependent upon the adrenals than the adrenals upon the brain, since the brain deteriorates progressively to death without the adrenals, while the adrenal whose connection with the brain has been broken by the division of its nerve supply will still produce sufficient adrenalin to support life.
From the strong affinity of the brain-cells for adrenalin which was manifested in our experiments we may strongly suspect that the Nissl substance is a volatile, extremely unstable combination of certain elements of the brain-cells and adrenalin, because the adrenals alone do not take the Nissl stain and the brain deprived of adrenalin also does not take Nissl stain. The consumption of the Nissl substance in the brain-cells is lessened or prevented by morphin, as is the output of adrenalin; and the consumption of the Nissl substance is also lessened or prevented by nitrous oxid. But morphin does not prevent the action of adrenalin injected into the circulation, hence the control of morphin over energy expenditure is exerted directly on the brain-cells. Apparently morphin and nitrous oxid both act
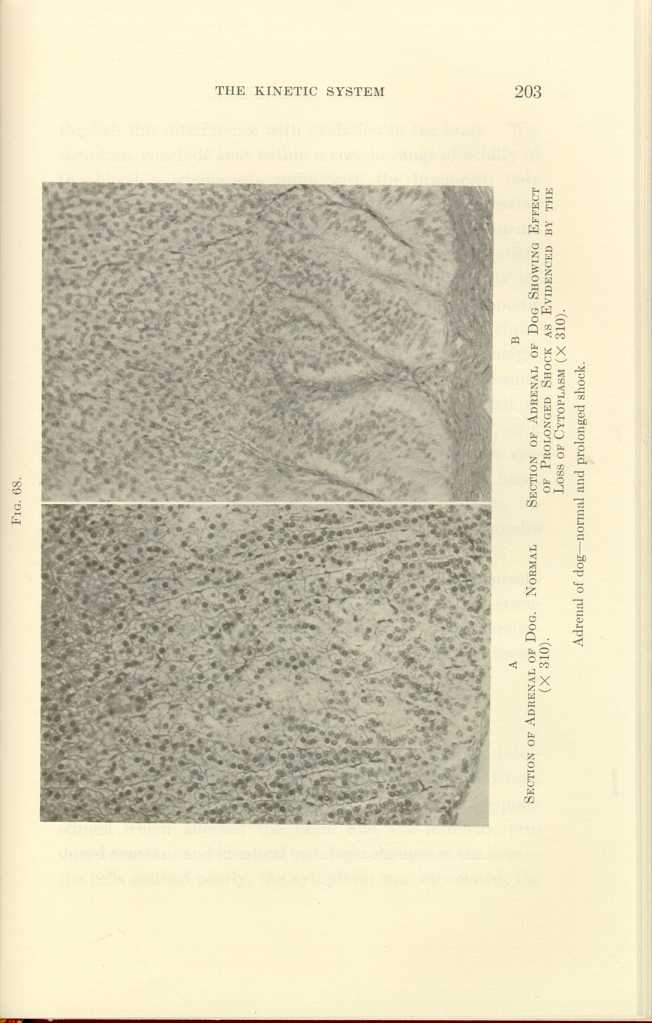
FIG. 68.
A: Section of Adrenal of Dog. Normal (x310).
B: Section of Adrenal of Dog Showing Effect of Prolonged Shock as
Evidenced by the Loss of Cytoplasm (x310).
Adrenal of dog—normal and prolonged shock.
[Description: Black-and-white photograph showing adrenal of dog under
various conditions.]
If this conclusion be well founded, we should find corroborative evidence in histologic changes in that great storehouse of potential energy, the liver, as a result of the application of each of the adequate stimuli which produced brain-cell and adrenal changes.
The Liver
Prolonged insomnia, prolonged physical exertion, infections, injections of toxins and of strychnin, rage and fear, physical injury under anesthesia, in fact, all the adequate stimuli which affected the brain and the adrenals, produced constant and identical histologic changes in the liver—the cells stained poorly, the cytoplasm was vacuolated, the
Are the histologic changes in the liver cells due to metabolism or toxic products, or are they "work" changes incident to the conversion of latent into kinetic energy? Are the brain, adrenals, and liver interdependent? The following facts establish the answers to these queries:
(1) The duration of life after excision of the liver is about the same as after adrenalectomy—approximately eighteen hours.
(2) The amount of glycogen in the liver was diminished in all the experiments showing brain-adrenal activity; and when the histologic changes were repaired, the normal amount of glycogen was again found.
(3) In crossed circulation experiments changes were found in the liver of the animal whose brain received the stimulus.
From these premises we must consider that the brain, the adrenals, and the liver are mutually dependent on one another for the conversion of latent into kinetic energy. Each is a vital organ, each equally vital. It may be said that excision of the brain may apparently cause death in less time than excision of the liver or adrenals, but this statement must be modified by our definition of death. If all the brain of an animal be removed by decapitation, its body may live on for at least eleven hours if its circulation be maintained by transfusion. An animal may live for weeks or months after excision of the cerebral hemispheres and the
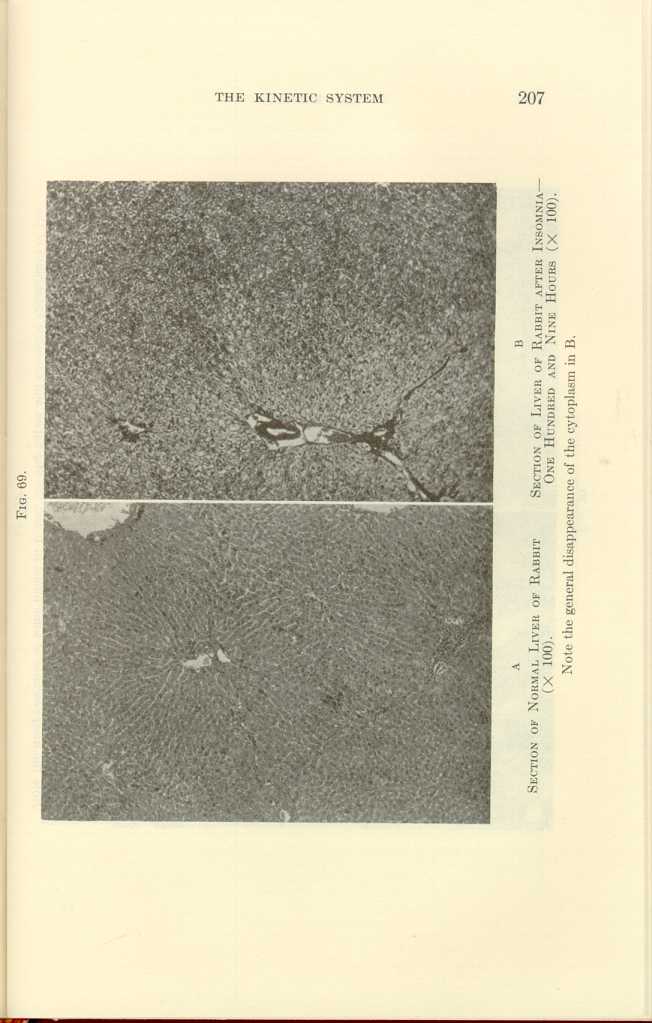
FIG. 69.
A: Section of Normal Liver of Rabbit (x100).
B: Section of Liver of Rabbit after Insomnia—One Hundred and Nine
Hours (x100).[k]
[Description: Black-and-white photograph showing liver of rabbit under
various conditions.]
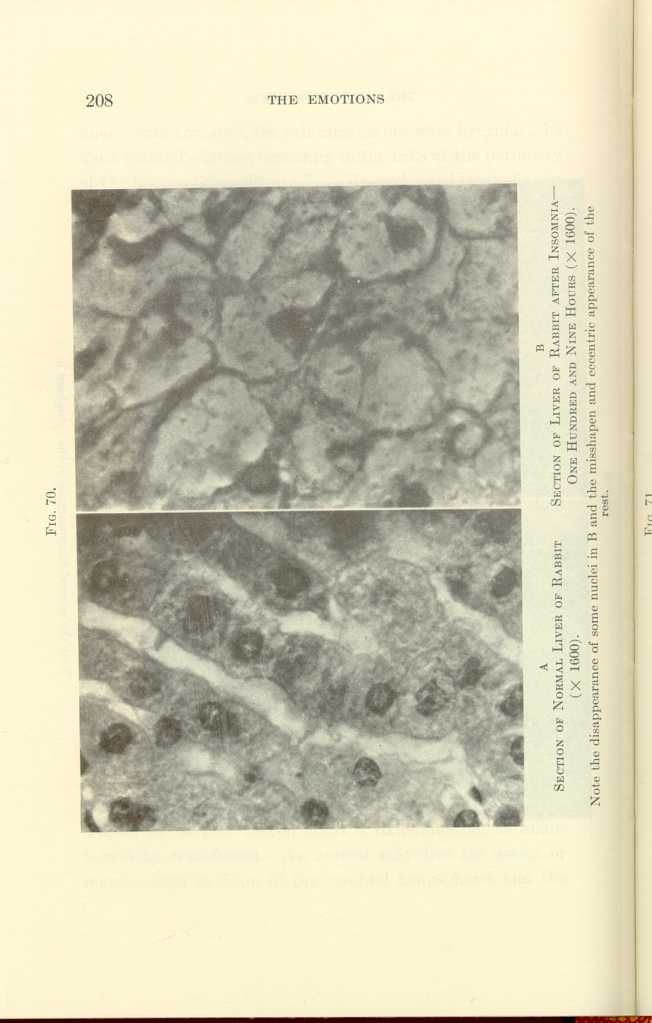
FIG. 70.
A: Section of Normal Liver of Rabbit (x1600).
B: Section of Liver of Rabbit after Insomnia—One Hundred and Nine
Hours (x1600).[l]
[Description: Black-and-white photograph showing liver of rabbit under
various conditions.]
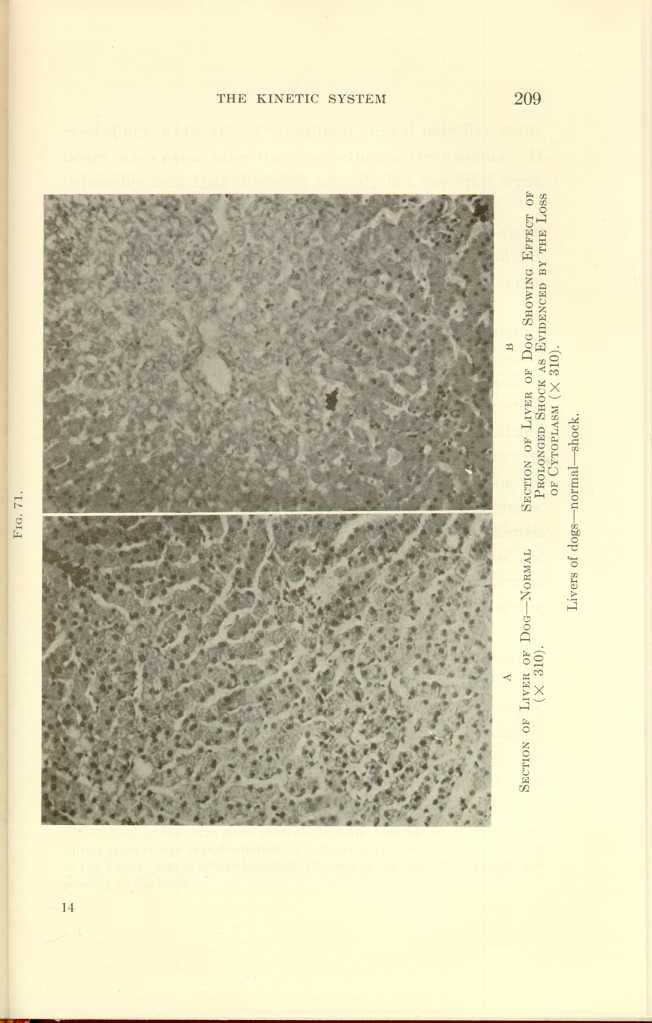
FIG. 71.
A: Section of Liver of Dog—Normal (x310).
B: Section of Liver of Dog Showing Effect of Prolonged Shock as
Evidenced by the Loss of Cytoplasm (x310).
Liver of dogs—normal—shock.
[Description: Black-and-white photograph showing liver of rabbit under
various conditions.]
In our research to discover whether any other organs should be included with the brain, the adrenals, and the liver in this mutually interdependent relation, we hit upon an experiment which throws light upon this problem.
Groups of rabbits were gently kept awake for one hundred hours by relays of students, an experiment which steadily withdrew energy but caused not the slightest physical or emotional injury to any of them; no drug, toxin, or other agent was given to them; they were given sufficient food and drink. In brief, the internal and external environments of these animals were kept otherwise normal excepting for the gentle stimuli which insured continued wakefulness. This protracted insomnia gradually exhausted the animals completely, some to the point of death even. Some of the survivors were killed immediately after the expiration of one hundred hours of wakefulness, others after varying intervals.
Histologic studies were made of every tissue and organ in the body. Three organs, the brain, the adrenals, and the liver, and these three only, showed histologic changes. In these three organs the histologic changes were marked, and were almost wholly repaired by one séance of sleep. In each instance these histologic changes were identical with those seen after physical exertion, emotions, toxins, etc.[4] It
With these three kinetic organs we may surely associate also the "furnace," the muscles, in which the energy provided by the brain, adrenals, and liver, plus oxygen, is fabricated into heat and motion.
Benedict, in his monumental work on metabolism, has demonstrated that in the normal state, at least, variations in the heart-beat parallel variations in metabolism. He and others have shown also that all the energy of the body, whether evidenced by heat or by motion, is produced in the muscles. In the muscles, then, we find the fourth vital link in the kinetic chain. The muscles move the body, circulate the blood, effect respiration, and govern the body temperature. They are the passive servants of the brain-adrenal-liver syndrome.
Neither the brain, the adrenals, the liver, nor the muscles, however, nor all of these together, have the power to change the rate of the expenditure of energy; to make possible the increased expenditure in adolescence, in pregnancy, in courting, and mating, in infections. No one of these organs, nor all of them together, can act as a pace-maker or sensitizer. The brain acts immediately in response to the stimuli of the moment; the adrenals respond instantly to the fickle brain and the effects of their actions are fleeting; the liver contains fuel only and cannot activate, and the muscles in turn act as the great furnace in which the final transformation into available energy is made.
The Thyroid
Another organ—the thyroid—has the special power of governing the rate of discharge of energy; in other words, the thyroid is the pace-maker. Unfortunately, the thyroid cannot be studied to advantage either functionally or histologically, for there is as yet no available test for thyroid secretion in the blood as there is for adrenalin, and thyroid activity is not attended by striking histologic changes. Therefore the only laboratory studies which have been satisfactory thus far are those by which the iodin content of the thyroid has been established. Iodin is stored in the colloid lacunæ of the thyroid and, in combination with certain proteins, is the active agent of the thyroid.
Beebe has shown that electric stimulation of the nerve supply of the thyroid diminishes the amount of iodin which it contains, and it is known that in the hyperactive thyroid in Graves' disease the iodin content is diminished. The meagerness of laboratory studies, however, is amply compensated by the observations which the surgeon has been able to make on a vast scale—observations which are as definite as are the results of laboratory experiments.
The brain-cells and the adrenals are securely, concealed from the eye of the clinician, hence the changes produced in them by different causes escape his notice, but the thyroid has always been closely scrutinized by him. The clinician knows that every one of the above-mentioned causes of increased brain-cell, adrenal, liver and muscle activity may cause an increase in the activity of both the normal or the enlarged thyroid; and lie knows only too well that in a given case of exophthalmic goiter the same stimuli which excite
The function of the thyroid in the kinetic chain is best evidenced, however, by its rôle in the production of fever. Fever results from the administration of thyroid extract alone in large doses. In the hyperactivity of the thyroid in exophthalmic goiter one sees a marked tendency to fever, in severe cases there is daily fever. In fact, in Graves' disease we find displayed to an extraordinary degree an exaggeration of the whole action of the kinetic mechanism.
We have stated that in acute Graves' disease there is a tendency to the production of spontaneous fever, and that there is a magnified diurnal variation in temperature which is due to an increased output of energy in even the normal reaction producing consciousness. In Graves' disease there is, therefore, a state of intensified consciousness, which is associated with low brain thresholds to all stimuli—both to stimuli that cause muscular action and to stimuli that cause fever. The intensity of the kinetic discharge is seen in the constant fine tremor. It is evident that the thresholds of the brain have been sensitized. In this hypersensitization we find the following strong evidence as to the identity of the various mechanisms for the production of fever. In the state of superlative sensitization which is seen in Graves' disease we find that the stimuli that produce muscular movement, the stimuli that produce emotional phenomena, and the stimuli that produce fever are as nearly as can be ascertained equally effective. Clinical evidence regarding this point is abundant, for in patients with Graves' disease we find that the three types of conversion of energy resulting from emotional stimulation, from infection stimulation, and
At will, then, through diminished, normal, or excessive administration of thyroid secretion, we may produce an adynamic, a normal, or an excessively dynamic state. By the thyroid influence, the brain thresholds are lowered and life becomes exquisite; without its influence the brain becomes a globe of relatively inert substance. Excessive doses of iodin alone cause most of the symptoms of Graves' disease. As we have stated, the active constituent of the thyroid is iodin in a special protein combination which is stored in the colloidal spaces. Hence one would not expect to find changes in the cells of the thyroid gland as a result of increased activity unless it be prolonged.
We have thus far considered the normal rôles played by the brain, the adrenals the liver, the muscles, and the thyroid in transforming latent into kinetic energy in the form of heat and motion as an adaptive response to environmental stimuli.
The argument may be strengthened, however, by the discussion of the effect of the impairment of any of these links in the kinetic chain upon the conversion of latent into kinetic energy.
Effect Upon the Output of Energy of Impaired or Lost Function of Each of the Several Links in the Kinetic Chain
(1) The Brain.—In cerebral softening we may find all the organs of the body comparatively healthy excepting the brain. As the brain is physically impaired it cannot normally stimulate other organs to the conversion of latent energy into heat or into motion, but, on the contrary, in these cases we find feeble muscular and intellectual power. I believe also that in patients with cerebral softening, infections such as pneumonia show a lower temperature range than in patients whose brains are normal.
(2) The Adrenals.—In such destructive lesions of the adrenals as Addison's disease one of the cardinal symptoms is a subnormal temperature and impaired muscular power. Animals upon whom double adrenalectomy has been performed show a striking fall in temperature, muscular weakness,—after adrenalectomy the animal may not be able to stand even,—and progressive chromatolysis.
(3) The Liver.—When the function of the liver is impaired by tumors, cirrhosis, or degeneration of the liver itself, then the entire energy of the body is correspondingly diminished. This diminution of energy is evidenced by muscular and mental weakness, by diminished response and by gradual loss of efficiency which finally reaches the state of asthenia.
(4) The Muscles.—It has been observed clinically that if the muscles are impaired by long disuse, or by a disease such
(5) The Thyroid.—In myxedema one of the cardinal symptoms is a persistently subnormal temperature and, though prone to infection, subjects of myxedema show but feeble febrile response and readily succumb. This clinical observation is strikingly confirmed by laboratory observations; normal rabbits subjected to fear showed a rise in temperature of from one to three degrees, while two rabbits whose thyroids had been previously removed and who had then been subjected to fright showed much less febrile response. Myxedema subjects show a loss of physical and mental energy which is proportional to the lack of thyroid. Deficiency in any of the organs of the kinetic chain causes alike loss of heat, loss of muscular and emotional action, of mental power, and of the power of combating infections—the negative evidence thus strongly supports the positive. By accumulating all the evidence we believe we are justified in associating the brain, the adrenals, the thyroid, the muscles, and the liver as vital links in the kinetic chain. Other organs play a rôle undoubtedly, though a minor one.
Studies in Hydrogen Ion Concentration in Activation of the Kinetic System
Having established the identity of some, at least, of the organs which constitute the kinetic chain, we endeavored to secure still further evidence regarding the energy-transforming function of these organs by making studies of the
H-ion concentration tests were made after the application of the adequate stimuli by which the function of the kinetic organs had been determined, and we studied also the effect upon the acidity of the blood of strychnin convulsions after destruction of the medulla; of deep narcotization with morphin before anesthesia; of deep narcotization with morphin after the H-ion concentration had already been increased by fear, by anger, by exertion, by injury under anesthesia, or by anesthesia alone.
The complete data of these experiments will be later reported in a monograph; here it is sufficient to state that anger, fear, injury, muscular exertion, inhalation anesthesia, strychnin, alcohol, in fact, all the stimuli which we had already found to produce histologic changes in the brain, the adrenals, and the liver-excepting bacterial toxins—caused increased H-ion concentration. Of striking significance is the fact that morphin alone caused no change in the H-ion concentration, while if administered before the application of a stimulus which by itself produced increased H-ion concentration, the action of that stimulus was neutralized or postponed. If, however, morphin was administered after increased acidity had been produced by any stimulus, or by inhalation anesthesia, then the time required for the restoration of the normal alkalinity was much prolonged, and in some instances the power of acid neutralization was permanently lost.
After excision of the liver, the normal H-ion concentration
In none of these cases was it determined whether the increased H-ion concentration was due to other causes of death or whether death was due to the increased acidity.
It is also significant that after the application of each of the adequate stimuli which increased the H-ion concentration of the blood in other parts of the body the blood from the adrenal vein showed a slight diminution in acidity, as, in most instances, did the blood from the hepatic vein also.
In fact, the H-ion concentration of the blood in the adrenal vein was less than in the blood of any other part of the circulation.
Kinetic Diseases
If our conclusions are sound, then in the kinetic system we find an explanation of many diseases, and having found the explanation, we may find new methods of combating them.
When the kinetic system is driven at an overwhelming rate of speed,—as by severe physical injury, by intense emotional excitation, by perforation of the intestines, by the pointing of an abscess into new territory, by the sudden onset of an infectious disease, by an overdose of strychnin,
The essential pathology of shock is identical whatever the cause. If, however, instead of an intense overwhelming activation, the kinetic system is continuously or intermittently overstimulated through a considerable period of time, as long as each of the links in the kinetic chain takes the strain equally the result will be excessive energy conversion, excessive work done; but usually, under stress, some one link in the chain is unable to take the strain and then the evenly balanced work of the several organs of the kinetic system is disturbed. If the brain cannot endure the strain, then neurasthenia, nerve exhaustion, or even insanity follows. If the thyroid cannot endure the strain, it undergoes hyperplasia, which in turn may result in a colloid goiter or in exophthalmic goiter. If the adrenals cannot endure the strain, cardiovascular disease may develop. If the liver cannot take the strain, then death from acute acidosis may follow, or if the neutralizing effect of the liver is only partially lost, then the acidity may cause Bright's disease. Overactivation of the kinetic system may cause glycosuria and diabetes.
Identical physical and functional changes in the organs of the kinetic system may result from intense continued stimulation from any of the following causes: Excessive physical labor, athletic exercise, worry or anxiety, intestinal auto-intoxication, chronic infections, such as oral sepsis, tonsillitis, and adenoids; chronic appendicitis, chronic cholecystitis,
From the foregoing statements we are able to understand the muscular weakness following fever; we can understand why the senile have neither muscular power nor strong febrile reaction; why long-continued infections produce pathologic changes in the organs constituting the kinetic chain; why the same pathologic changes result from various forms of activation of the kinetic system. In this hypothesis we find a reason why cardiovascular disease may be caused by chronic infection, by auto-intoxication, by overwork, or by emotional excitation. We now see that the reason why we find so much difficulty in differentiating the numerous acute infections from each other is because they play upon the same kinetic chain. Our postulate harmonizes the pathologic democracy of the kinetic organs, for it explains not only why, in many diseases, the pathologic changes in these organs are identical, but why the same changes are seen as the result of emotional strain and overwork. We can thus understand how either emotional strain or acute or chronic infection may cause either exophthalmic goiter or cardiovascular disease; how chronic intestinal stasis with the resultant absorption of toxins may cause cardiovascular disease, neurasthenia, or goiter. Here is found an explanation of the phenomena of shock, whether the shock be the result of toxins, of infection, of foreign proteins, of anaphylaxis, of psychic stimuli, or of a surgical operation with its combination of both psychic and traumatic elements.
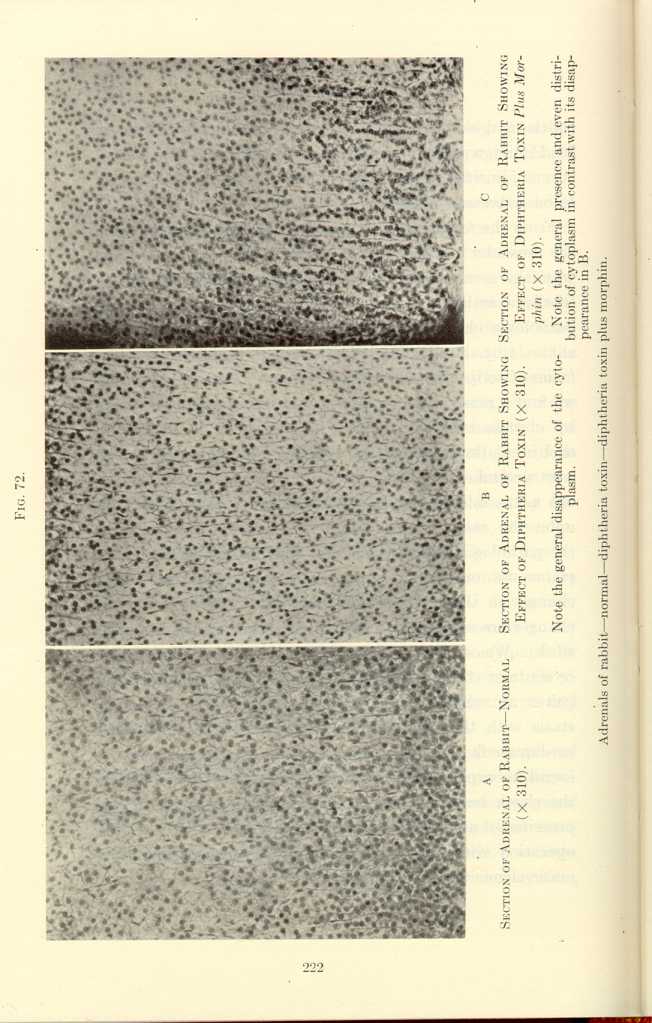
FIG. 72[m]
A: Section of Adrenal of Rabbit—Normal (x310).
B: Section of Adrenal of Rabbit Showing Effect of Diptheria Toxin
(x310).[n]
C: Section of Adrenal of Rabbit Showing Effect of Diptheria Toxin
Plus Morphin (x310).[o]
[Description: Black-and-white photographs showing adrenal of rabbit under
various conditions.]
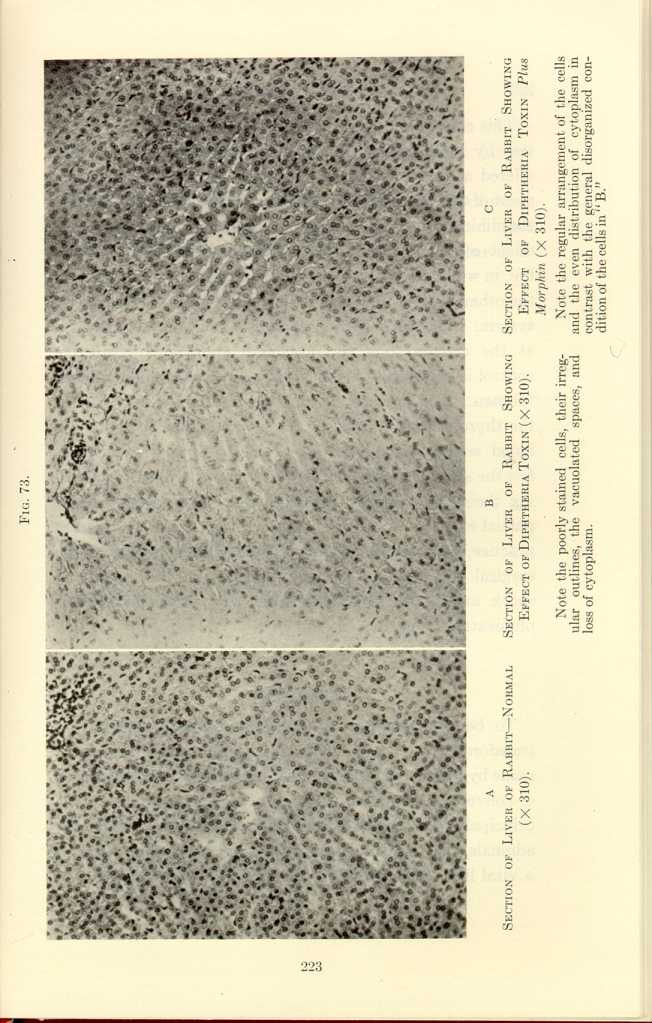
FIG. 73.
A: Section of Adrenal of Rabbit—Normal (x310).
B: Section of Adrenal of Rabbit Showing Effect of Diptheria Toxin
(x310).[p]
C: Section of Adrenal of Rabbit Showing Effect of Diptheria Toxin
Plus Morphin (x310).[q]
[Description: Black-and-white photographs showing adrenal of rabbit under
various conditions.]
This conception of the kinetic system has stood a crucial test by making possible the shockless operation. It has offered a plausible explanation of the cause and the treatment of Graves' disease. Will the kinetic theory stand also the clinical test of controlling that protean disease bred in the midst of the stress of our present-day life? Present-day life, in which one must ever have one hand on the sword and the other on the throttle, is a constant stimulus of the kinetic system. The force of these kinetic stimuli may be lessened at the cerebral link by intelligent control—a protective control is empirically attained by many of the most successful men. The force of the kinetic stimuli may be broken at the thyroid link by dividing the nerve supply, reducing the blood supply, or by partial excision; or if the adrenals feel the strain, the stimulating force may be broken by dividing their nerve supply, reducing the blood supply, or by partial excision. No theory is worth more than its yield in practice, but already we have the shockless operation, the surgical treatment of Graves' disease, and the control of shock and of the acute infections by overwhelming morphinization (Figs. 62, 72, and 73).
Conclusions
To become adapted to their environment animals are transformers of energy. This adaptation to environment is made by means of a system of organs evolved for the purpose of converting potential energy into heat and motion. The principal organs and tissues of this system are the brain, the adrenals, the thyroid, the muscles, and the liver. Each is a vital link, each plays its particular rôle, and one cannot
In this conception we find a possible explanation of many diseases one which may point the way to new and more effective therapeutic measures than those now at our command.
Address delivered before the New York State Medical Society, April 28, 1914, to which has been added a further note regarding studies of hydrogen ion concentration in the blood.
From H. K. Cushing Laboratory of Experimental Medicine, Western Reserve University, Cleveland.
We use the terms "heat" and "muscular action" in the popular sense, though physicists use them to designate one and the same kind of energy.
Further studies have given evidence that the elimination of the acids resulting from energy-transformation as well as the conversion of energy stored in the kinetic organs causes histologic changes in the liver, the adrenals, and possibly in the brain.
There is a general loss of cyoplasm and but faint traces of most of the Purkinje cells. This photograph presents striking evidence of the direct dependence of the brain upon the liver.
The disastrous effect of the withdrawal of adrenalin from teh system is marked by the depleted cells. Every Purkinje cell shows signs of exhaustion, some are completely disintegrated. This and the following photomicrograph (C) make evident the dependence of the brain upon the adrenals.
The remarkable activating effect of adrenalin is strikingly shown by teh intense hyperchromatism.
The protective effect of morphin is illustrated by the large percentage of hyperchromatic cells.
The patient's brain received neither traumatic nor psychic stimuli from the time she was anesthetized in bed until she returned again from the operating-room. Her pulse-rate fell slightly during the operation. On the other hand, the psychic strain undergone by the patient's sister while the operation was being performed caused her pulse-rate to rise to 124.
—The patient, a foreigner, was brought to the operating-room from the accident ward. Pulse and temperature normal. When he found himself in the operating-room he was greatly disturbed. It was impossible to make him understand that his leg was not to be amputated but only a plaster cast applied. Under this stimulus his pulse rose to 150 and he soon developed a temperature of 101.2° F.
Note the eccentric and irregularly shaped nuclei and the generally disorganized appearance of the cells in B.
Note the disappearance of some nuclei in B and the misshapen and eccentric appearance of the rest.
Note the general presence and even distribution of cytoplasm in contrast with its disappearance in B.
| The Origin and Nature of the Emotions: Miscellaneous Papers | ||